

The primary σ factor in Escherichia coli can access the transcription elongation complex from solution in vivo
- PMID: 26371553
- PMCID: PMC4604602
- DOI: 10.7554/eLife.10514
The primary σ factor in Escherichia coli can access the transcription elongation complex from solution in vivo
Abstract
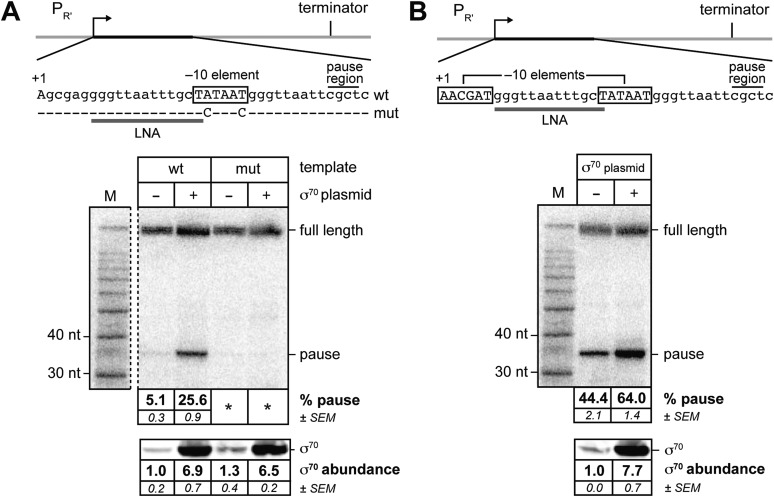
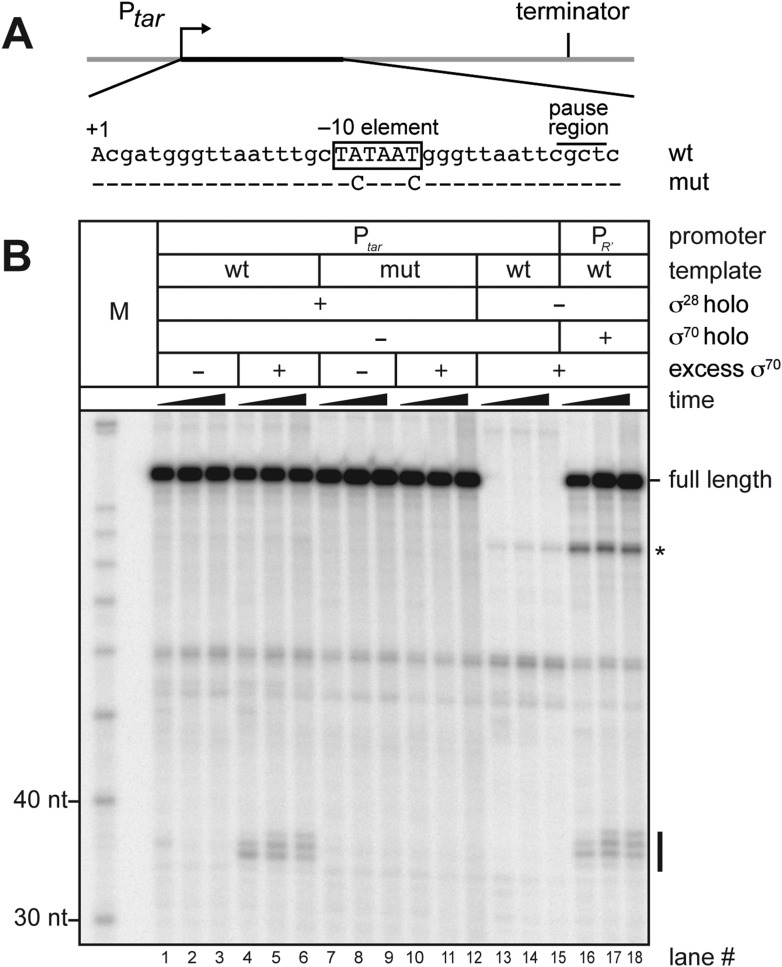
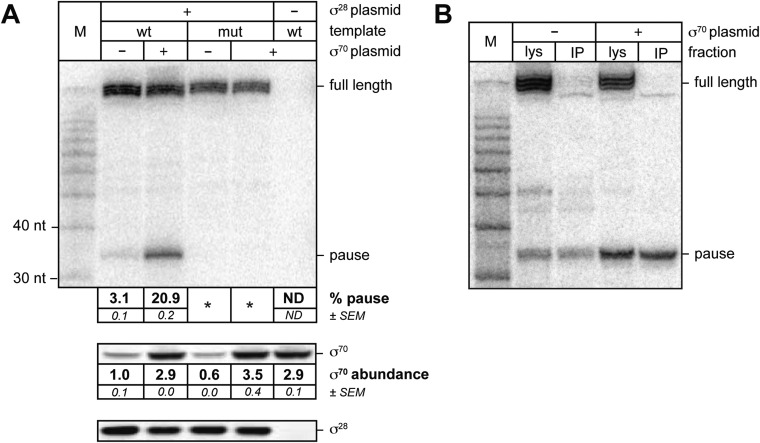
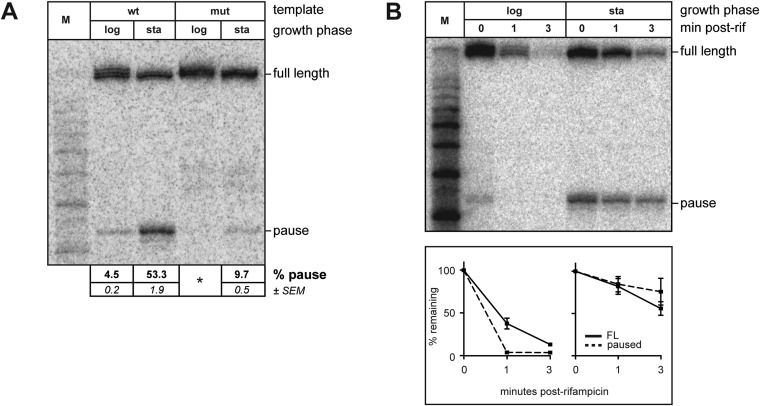
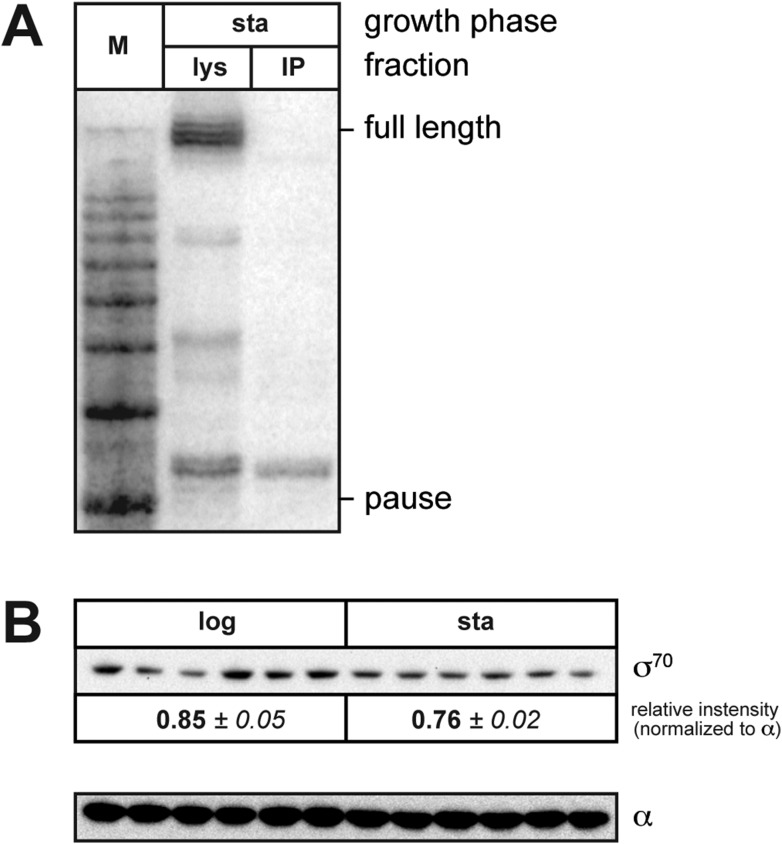
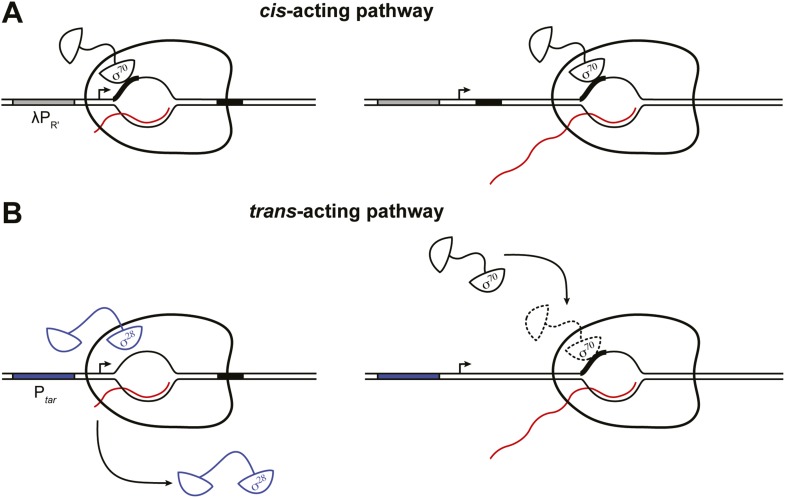
The σ subunit of bacterial RNA polymerase (RNAP) confers on the enzyme the ability to initiate promoter-specific transcription. Although σ factors are generally classified as initiation factors, σ can also remain associated with, and modulate the behavior of, RNAP during elongation. Here we establish that the primary σ factor in Escherichia coli, σ(70), can function as an elongation factor in vivo by loading directly onto the transcription elongation complex (TEC) in trans. We demonstrate that σ(70) can bind in trans to TECs that emanate from either a σ(70)-dependent promoter or a promoter that is controlled by an alternative σ factor. We further demonstrate that binding of σ(70) to the TEC in trans can have a particularly large impact on the dynamics of transcription elongation during stationary phase. Our findings establish a mechanism whereby the primary σ factor can exert direct effects on the composition of the entire transcriptome, not just that portion that is produced under the control of σ(70)-dependent promoters.
Keywords: E. coli; RNA polymerase; bacterial transcription; chromosomes; elongation factor; genes; infectious disease; microbiology; sigma factor; transcription pausing.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
