

Luteinizing hormone induces ovulation via tumor necrosis factor α-dependent increases in prostaglandin F2α in a nonmammalian vertebrate
- PMID: 26374476
- PMCID: PMC4570979
- DOI: 10.1038/srep14210
Luteinizing hormone induces ovulation via tumor necrosis factor α-dependent increases in prostaglandin F2α in a nonmammalian vertebrate
Abstract
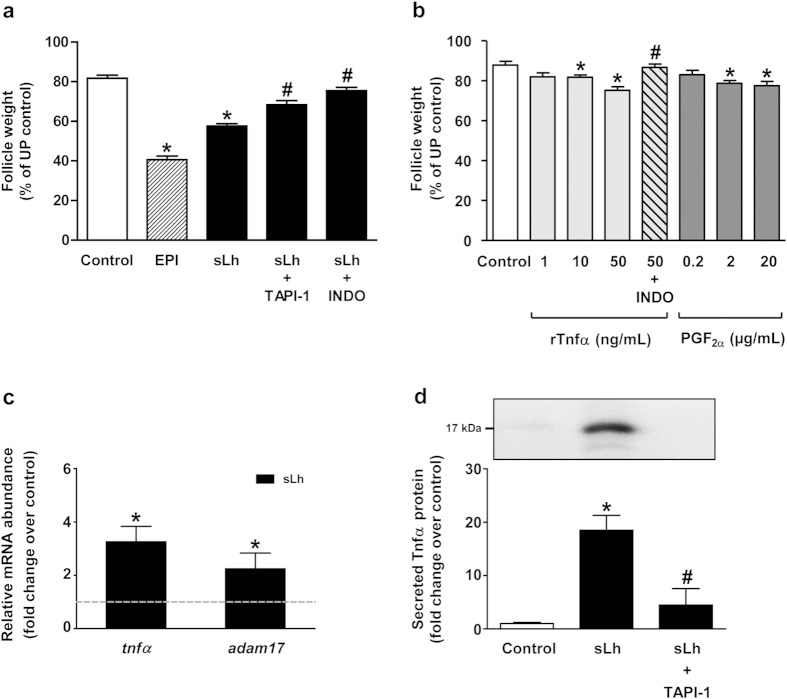
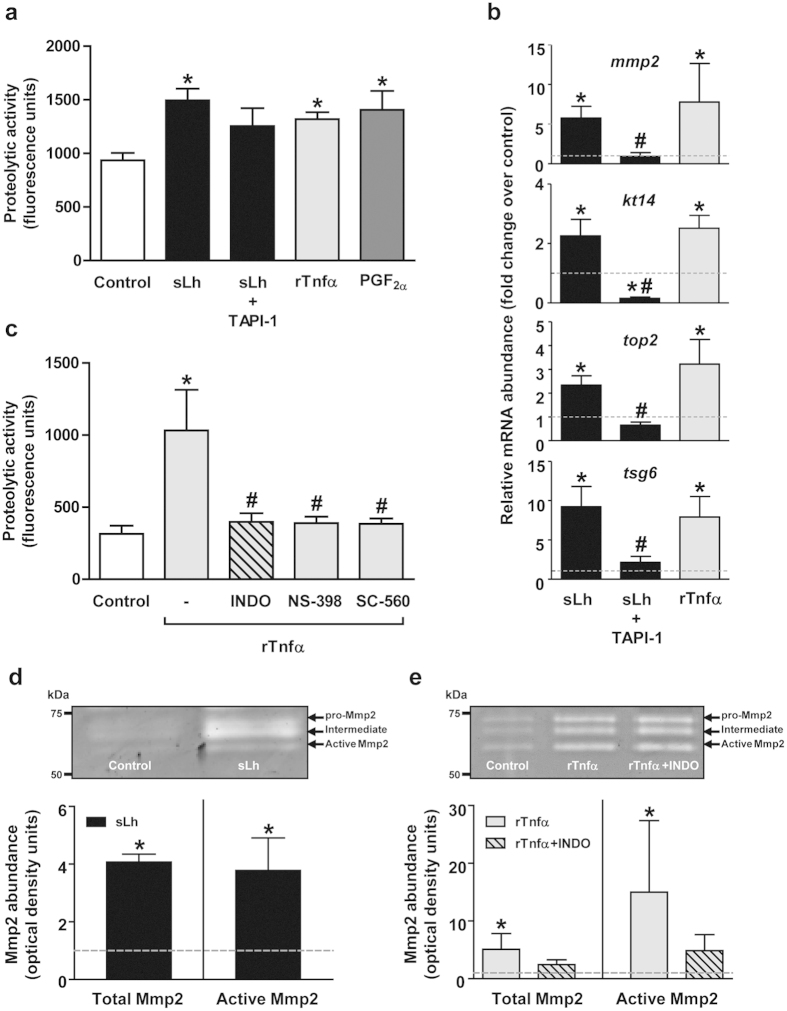
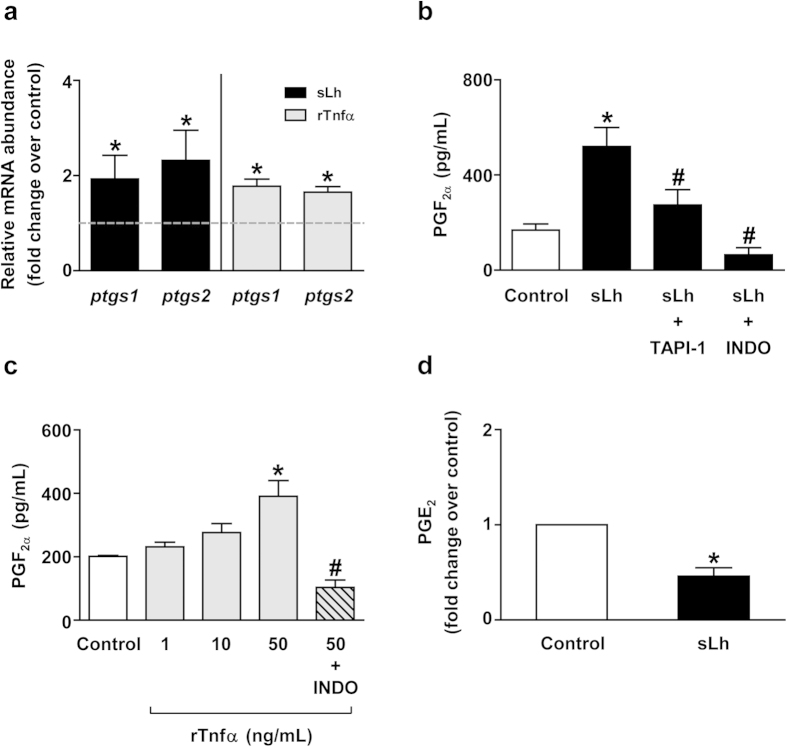
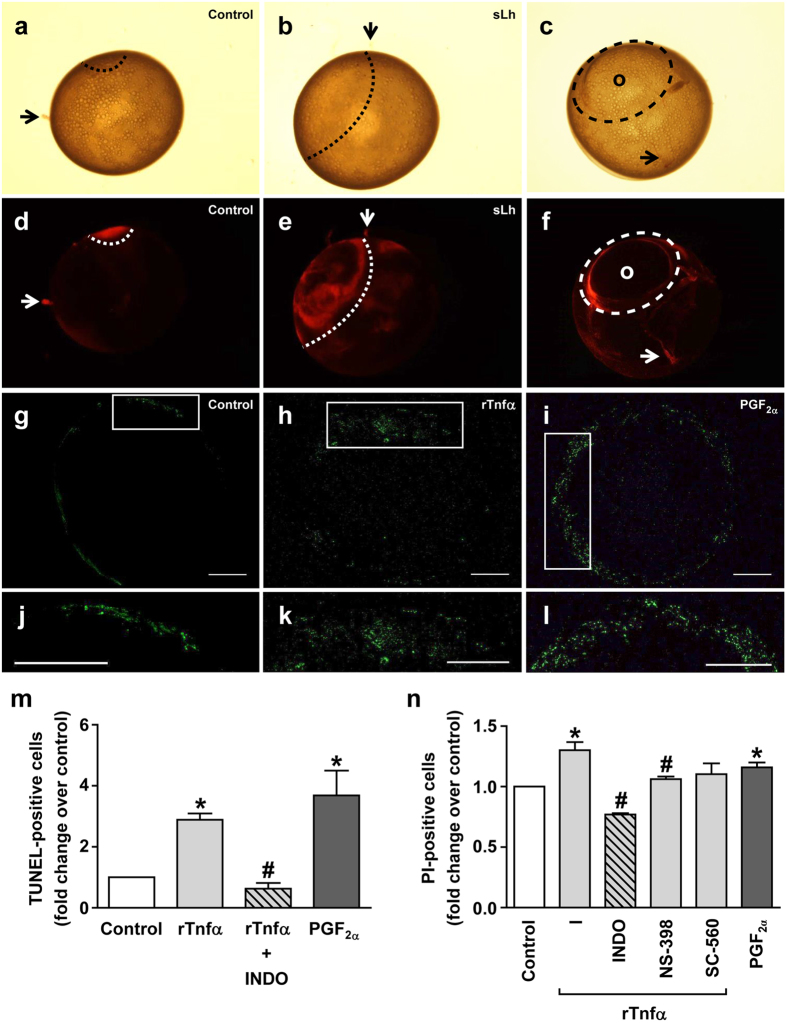
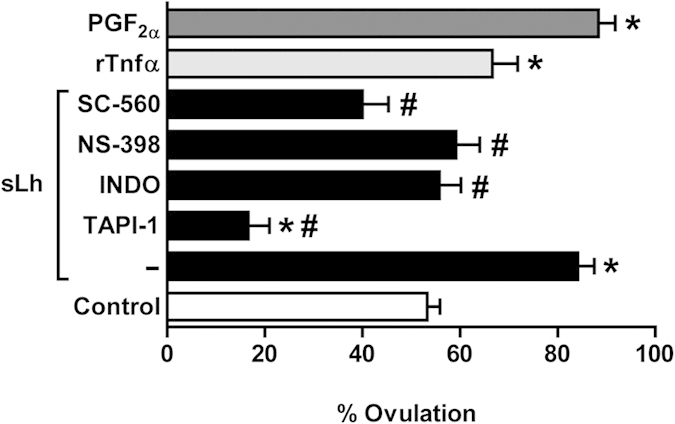
Ovulation is induced by the preovulatory surge of luteinizing hormone (LH) that acts on the ovary and triggers the rupture of the preovulatory ovarian follicle by stimulating proteolysis and apoptosis in the follicle wall, causing the release of the mature oocyte. The pro-inflammatory cytokine tumor necrosis factor α (TNFα) and prostaglandin (PG) F2α (PGF2α) are involved in the control of ovulation but their role mediating the pro-ovulatory actions of LH is not well established. Here we show that Lh induces PGF2α synthesis through its stimulation of Tnfα production in trout, a primitive teleost fish. Recombinant trout Tnfα (rTnfα) and PGF2α recapitulate the stimulatory in vitro effects of salmon Lh (sLh) on contraction, proteolysis and loss of cell viability in the preovulatory follicle wall and, finally, ovulation. Furthermore, all pro-ovulatory actions of sLh are blocked by inhibition of Tnfα secretion or PG synthesis and all actions of rTnfα are blocked by PG synthesis inhibitors. Therefore, we provide evidence that the Tnfα-dependent increase in PGF2α production is necessary for the pro-ovulatory actions of Lh. The results from this study shed light onto the mechanisms underlying the pro-ovulatory actions of LH in vertebrates and may prove important in clinical assessments of female infertility.
Figures
References
-
- Espey L. L. & Richards J. S. Ovulation in The Physiology of Reproduction (ed. Neil J. D.) 425–474 (Academic Press, 2006).
-
- Rao C. V. & Lei Z. M. Consequences of targeted inactivation of LH receptors. Mol Cell Endocrinol 187, 57–67 (2002). - PubMed
-
- Curry T. E. J. & Smith M. F. Impact of extracellular matrix remodeling on ovulation and the folliculo-luteal transition. Semin Reprod Med 24, 228–241 (2006). - PubMed
-
- Reich R. et al. Preovulatory changes in ovarian expression of collagenases and tissue metalloproteinase inhibitor messenger ribonucleic acid: role of eicosanoids. Endocrinology 129, 1869–1875 (1991). - PubMed
-
- Gottsch M. L., Van Kirk E. A. & Murdoch W. J. Tumour necrosis factor alpha up-regulates matrix metalloproteinase-2 activity in periovulatory ovine follicles: metamorphic and endocrine implications. Reprod Fertil Dev 12, 75–80 (2000). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
