

Conditional depletion of intellectual disability and Parkinsonism candidate gene ATP6AP2 in fly and mouse induces cognitive impairment and neurodegeneration
- PMID: 26376863
- PMCID: PMC4634377
- DOI: 10.1093/hmg/ddv380
Conditional depletion of intellectual disability and Parkinsonism candidate gene ATP6AP2 in fly and mouse induces cognitive impairment and neurodegeneration
Abstract
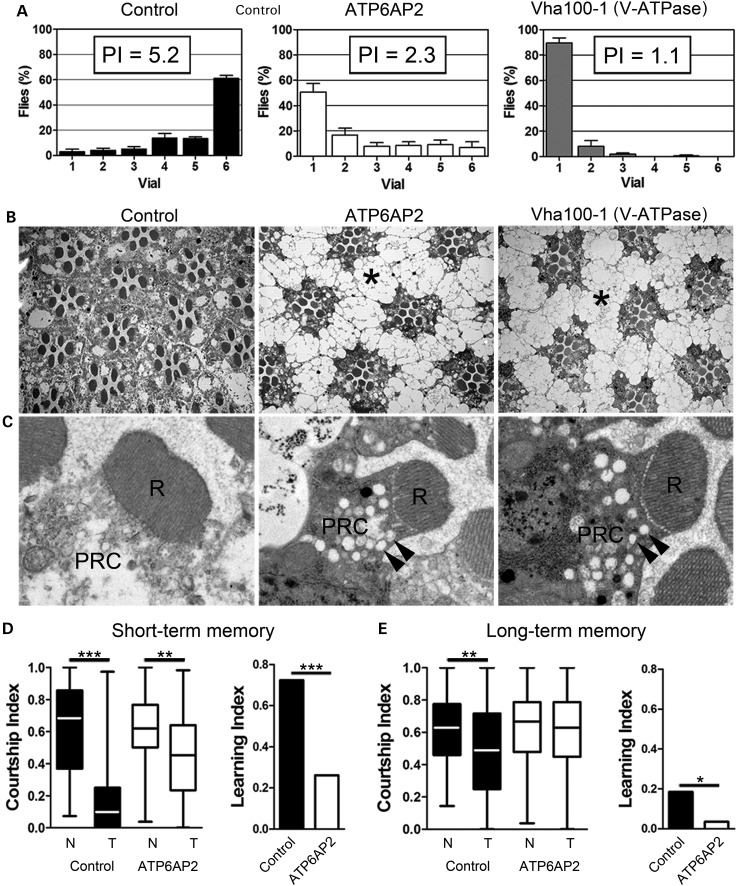
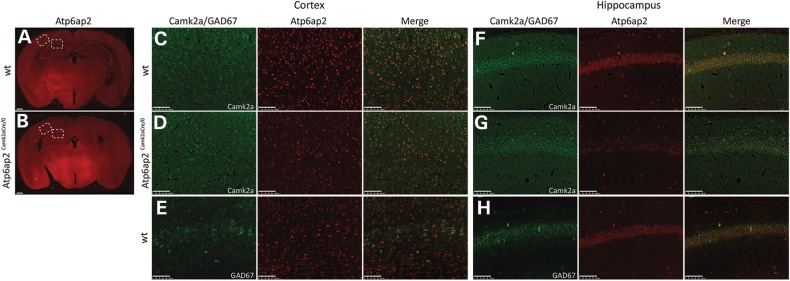
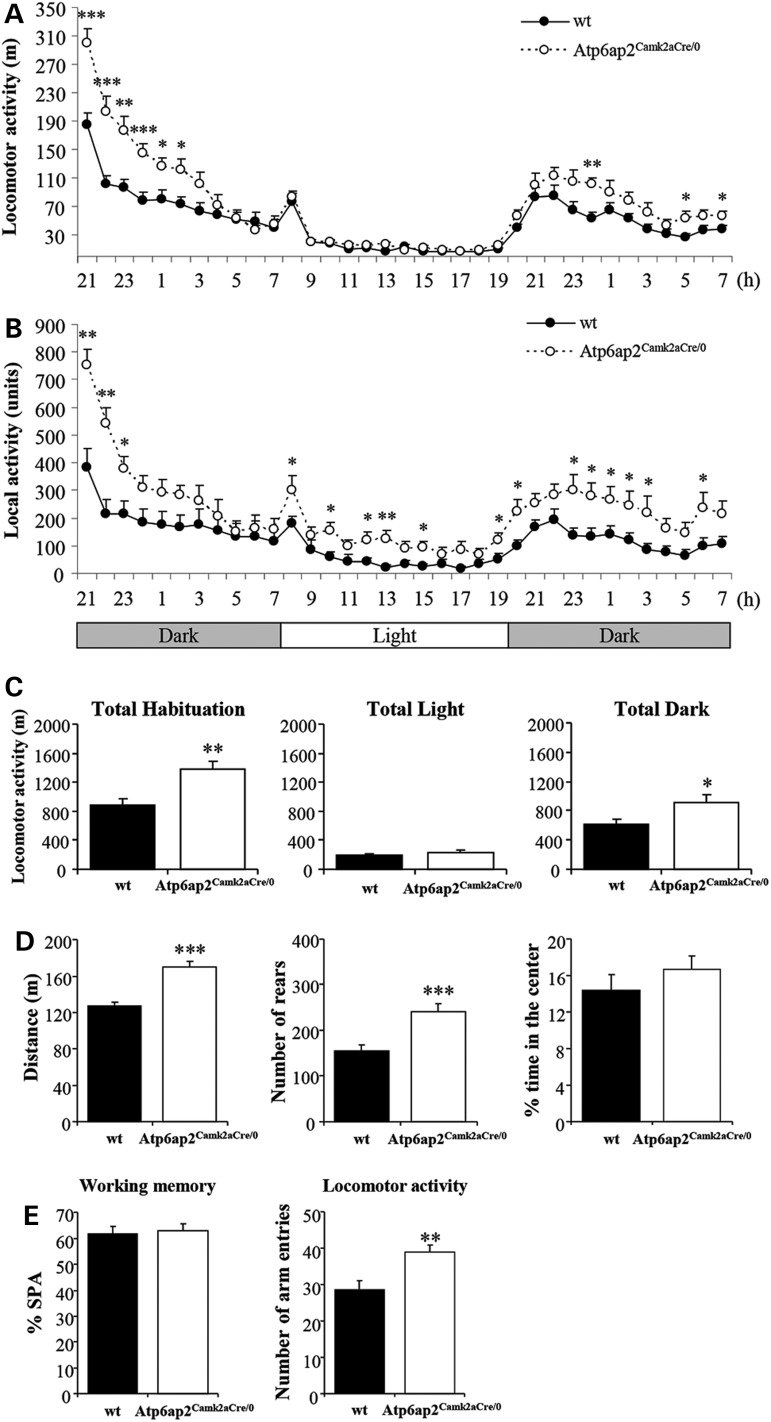
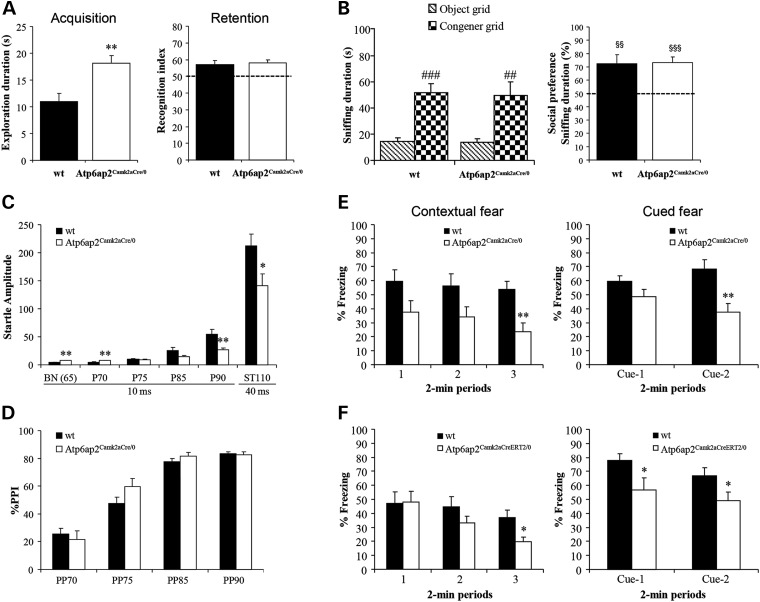
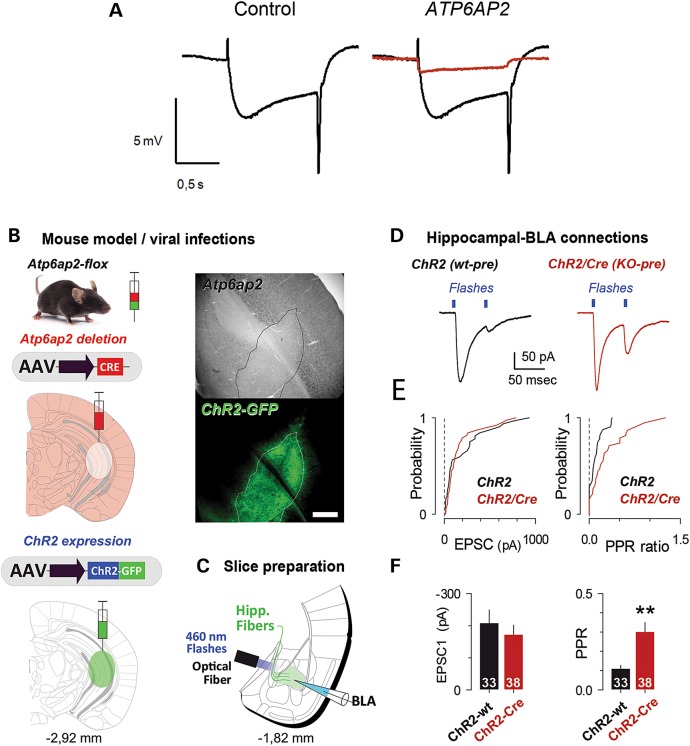
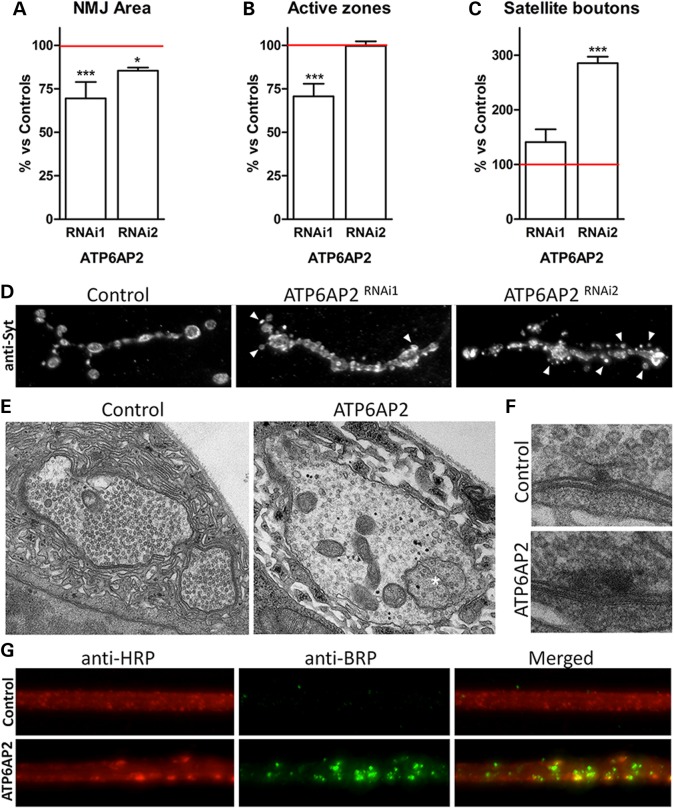
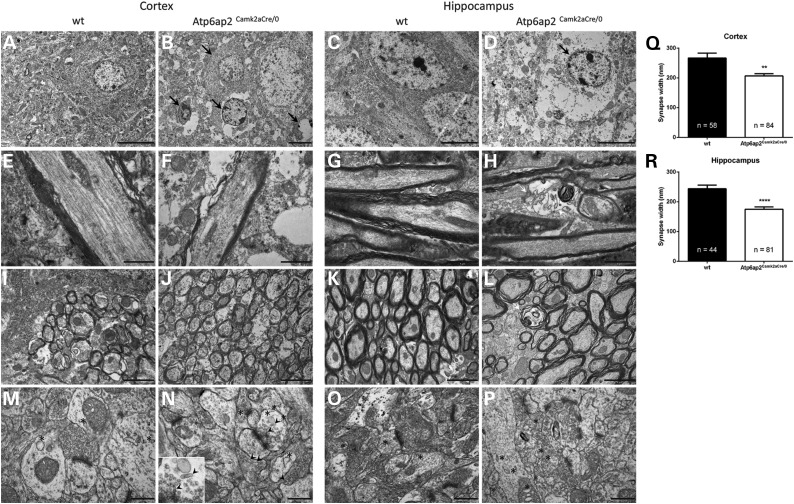
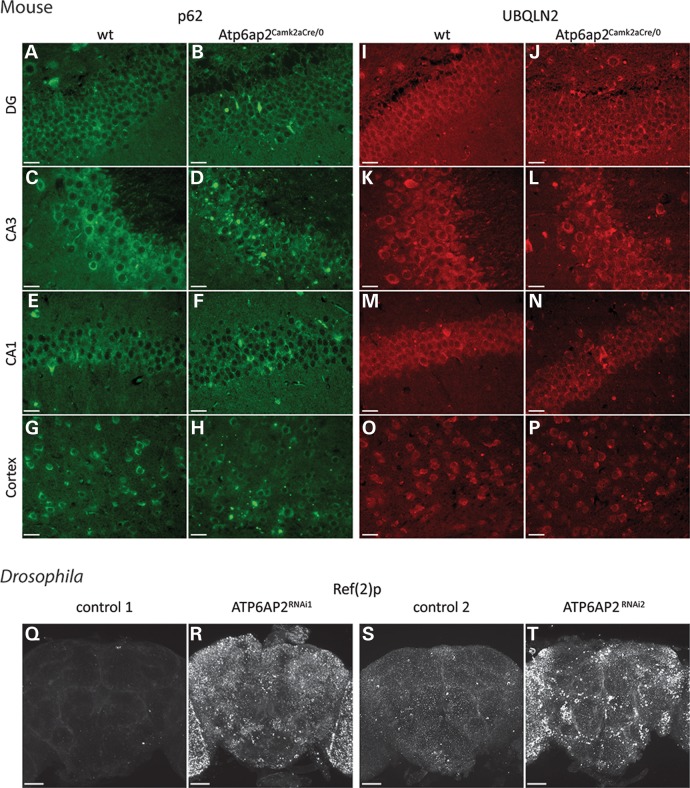
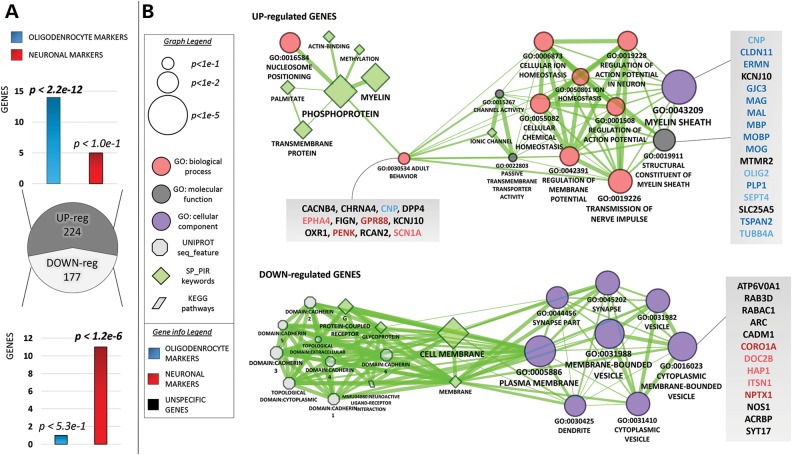
ATP6AP2, an essential accessory component of the vacuolar H+ ATPase (V-ATPase), has been associated with intellectual disability (ID) and Parkinsonism. ATP6AP2 has been implicated in several signalling pathways; however, little is known regarding its role in the nervous system. To decipher its function in behaviour and cognition, we generated and characterized conditional knockdowns of ATP6AP2 in the nervous system of Drosophila and mouse models. In Drosophila, ATP6AP2 knockdown induced defective phototaxis and vacuolated photoreceptor neurons and pigment cells when depleted in eyes and altered short- and long-term memory when depleted in the mushroom body. In mouse, conditional Atp6ap2 deletion in glutamatergic neurons (Atp6ap2(Camk2aCre/0) mice) caused increased spontaneous locomotor activity and altered fear memory. Both Drosophila ATP6AP2 knockdown and Atp6ap2(Camk2aCre/0) mice presented with presynaptic transmission defects, and with an abnormal number and morphology of synapses. In addition, Atp6ap2(Camk2aCre/0) mice showed autophagy defects that led to axonal and neuronal degeneration in the cortex and hippocampus. Surprisingly, axon myelination was affected in our mutant mice, and axonal transport alterations were observed in Drosophila. In accordance with the identified phenotypes across species, genome-wide transcriptome profiling of Atp6ap2(Camk2aCre/0) mouse hippocampi revealed dysregulation of genes involved in myelination, action potential, membrane-bound vesicles and motor behaviour. In summary, ATP6AP2 disruption in mouse and fly leads to cognitive impairment and neurodegeneration, mimicking aspects of the neuropathology associated with ATP6AP2 mutations in humans. Our results identify ATP6AP2 as an essential gene for the nervous system.
© The Author 2015. Published by Oxford University Press.
Figures
References
-
- Hedera P., Alvarado D., Beydoun A., Fink J.K. (2002) Novel mental retardation-epilepsy syndrome linked to Xp21.1-p11.4. Ann. Neurol., 51, 45–50. - PubMed
-
- Ramser J., Abidi F.E., Burckle C.A., Lenski C., Toriello H., Wen G., Lubs H.A., Engert S., Stevenson R.E., Meindl A. et al. (2005) A unique exonic splice enhancer mutation in a family with X-linked mental retardation and epilepsy points to a novel role of the renin receptor. Hum. Mol. Genet., 14, 1019–1027. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
