

Prox1 Regulates the Subtype-Specific Development of Caudal Ganglionic Eminence-Derived GABAergic Cortical Interneurons
- PMID: 26377473
- PMCID: PMC4571608
- DOI: 10.1523/JNEUROSCI.1164-15.2015
Prox1 Regulates the Subtype-Specific Development of Caudal Ganglionic Eminence-Derived GABAergic Cortical Interneurons
Abstract
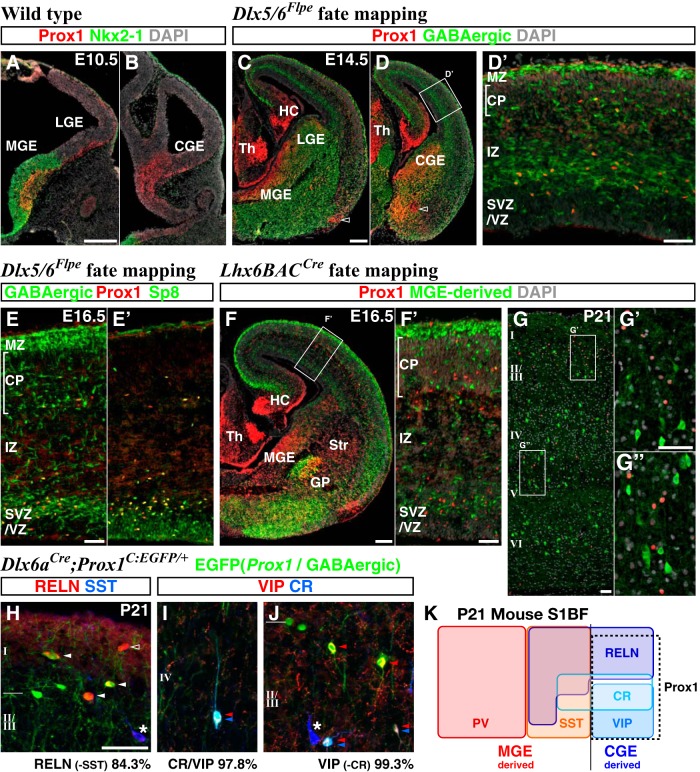
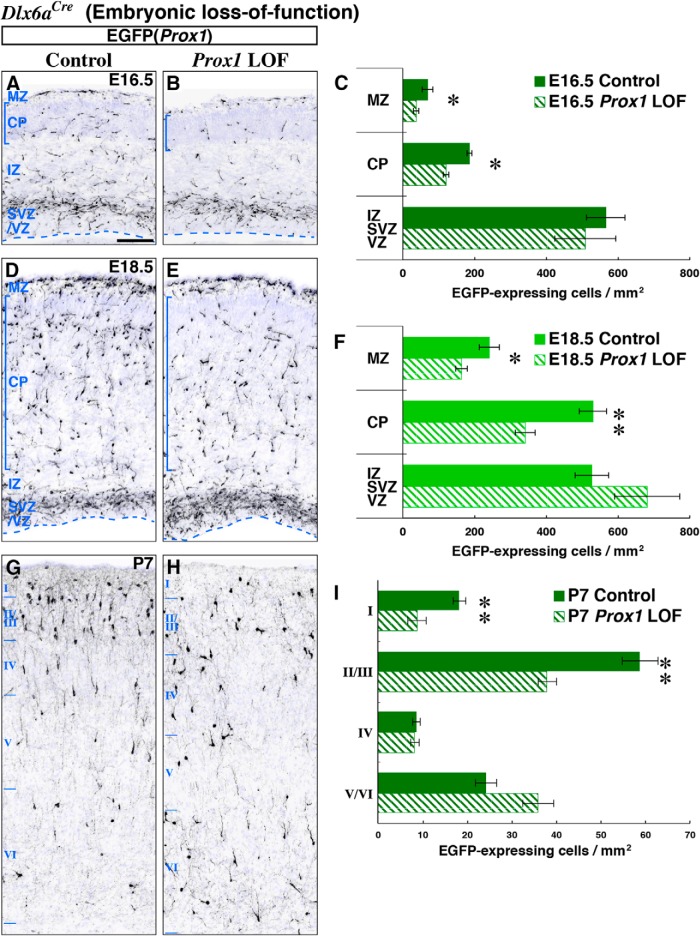
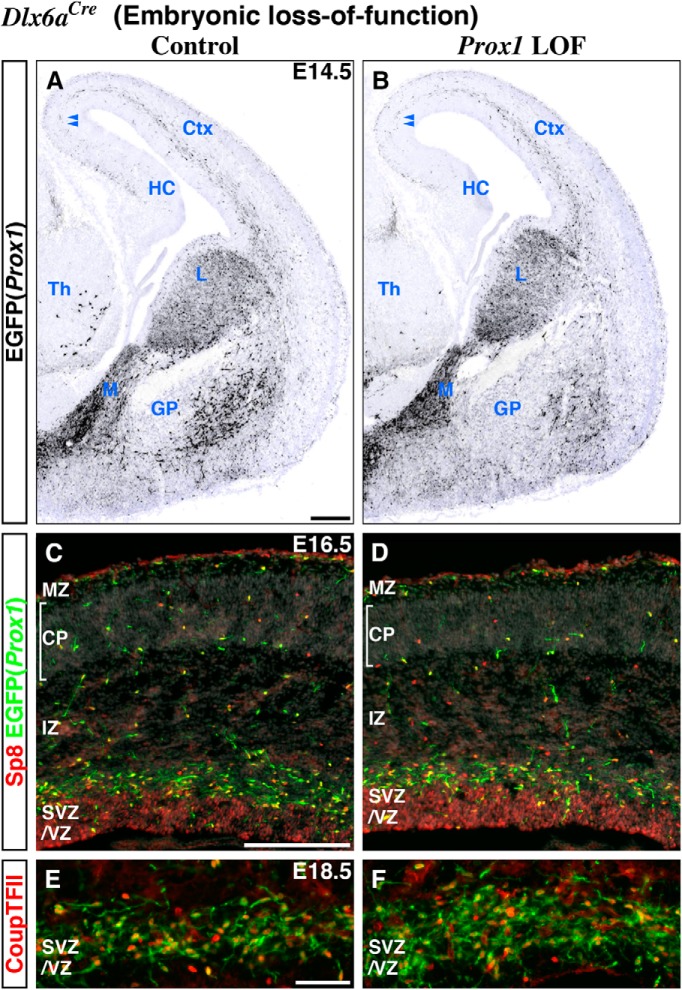
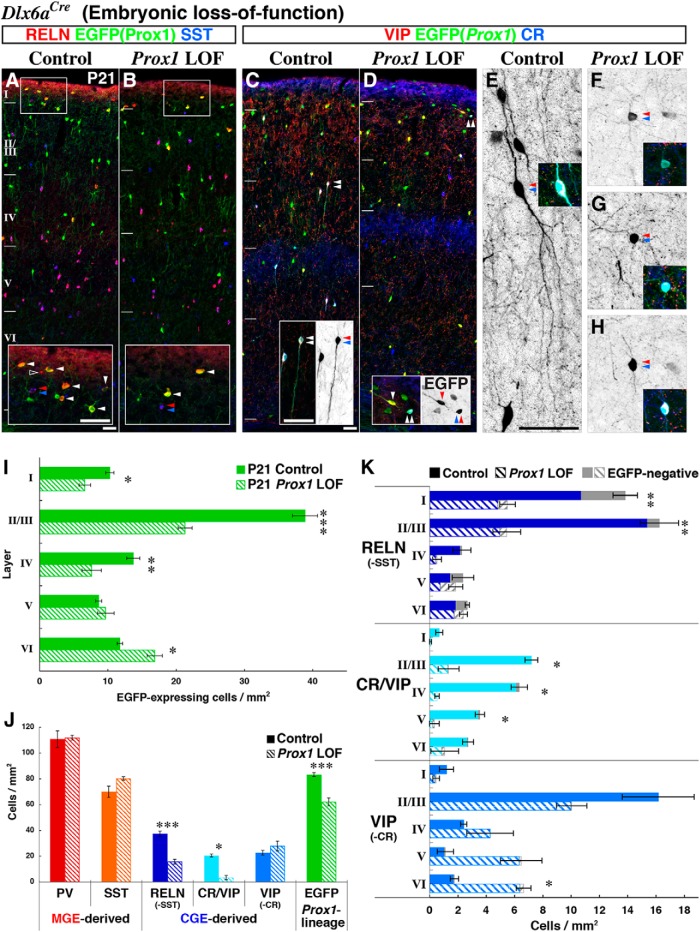
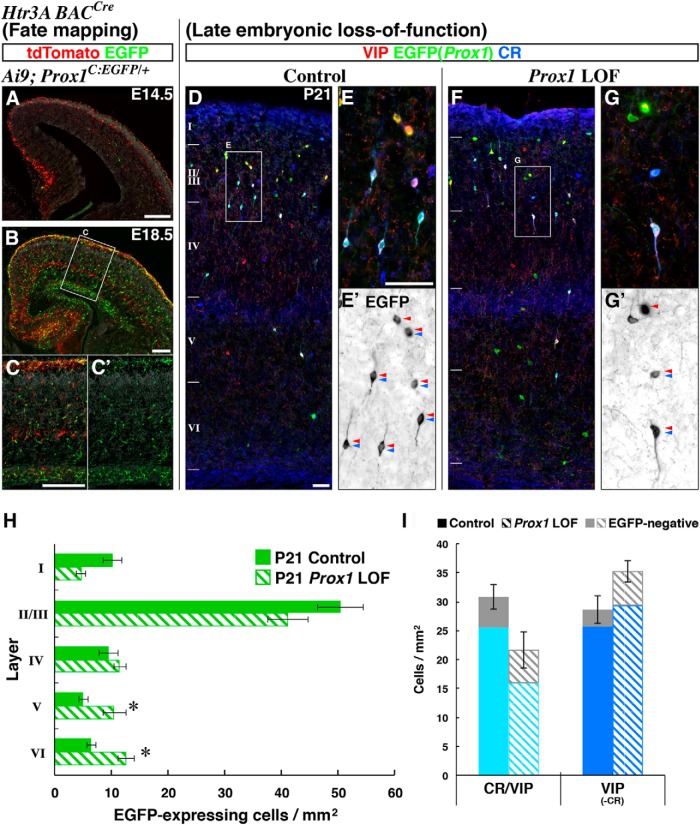
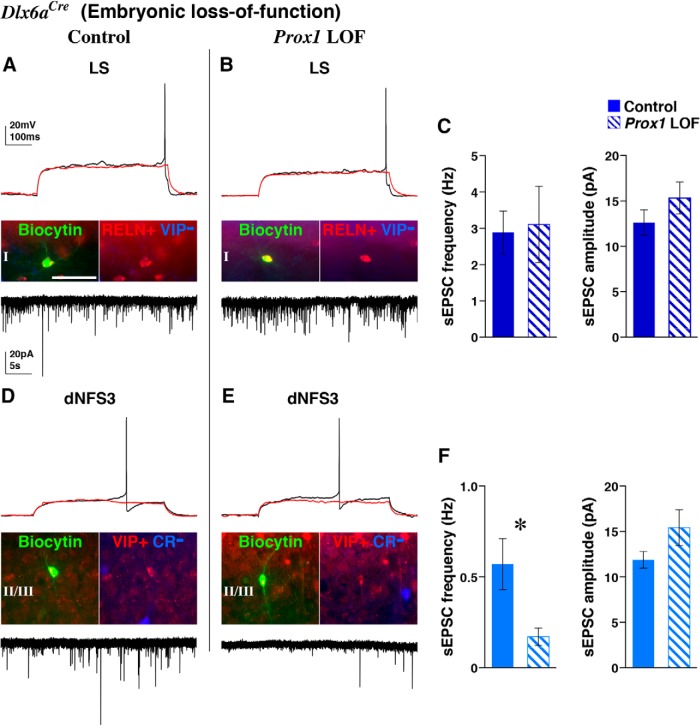
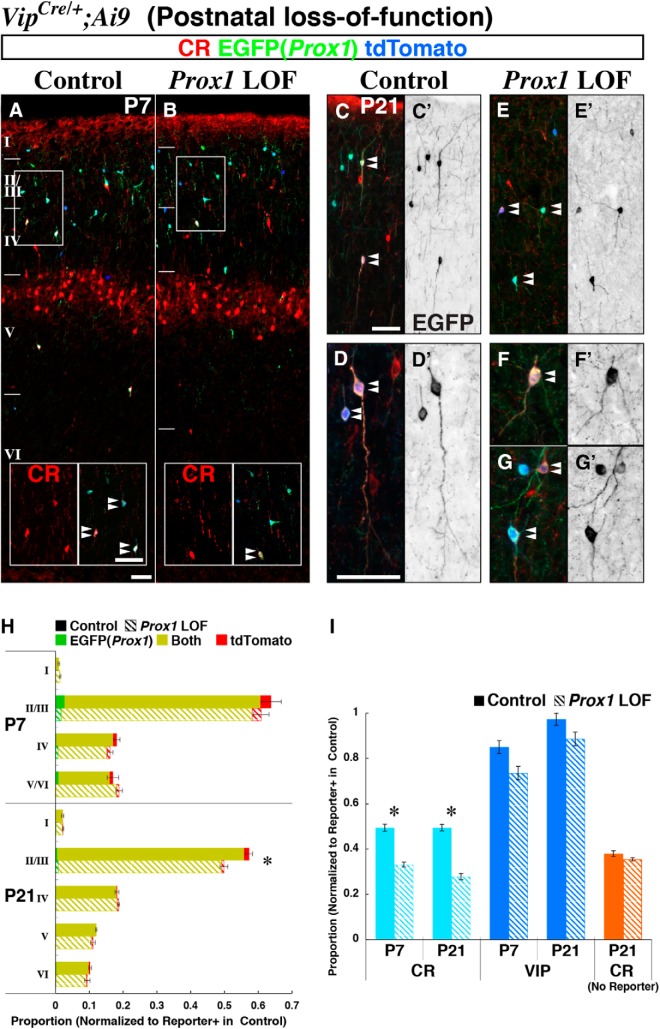
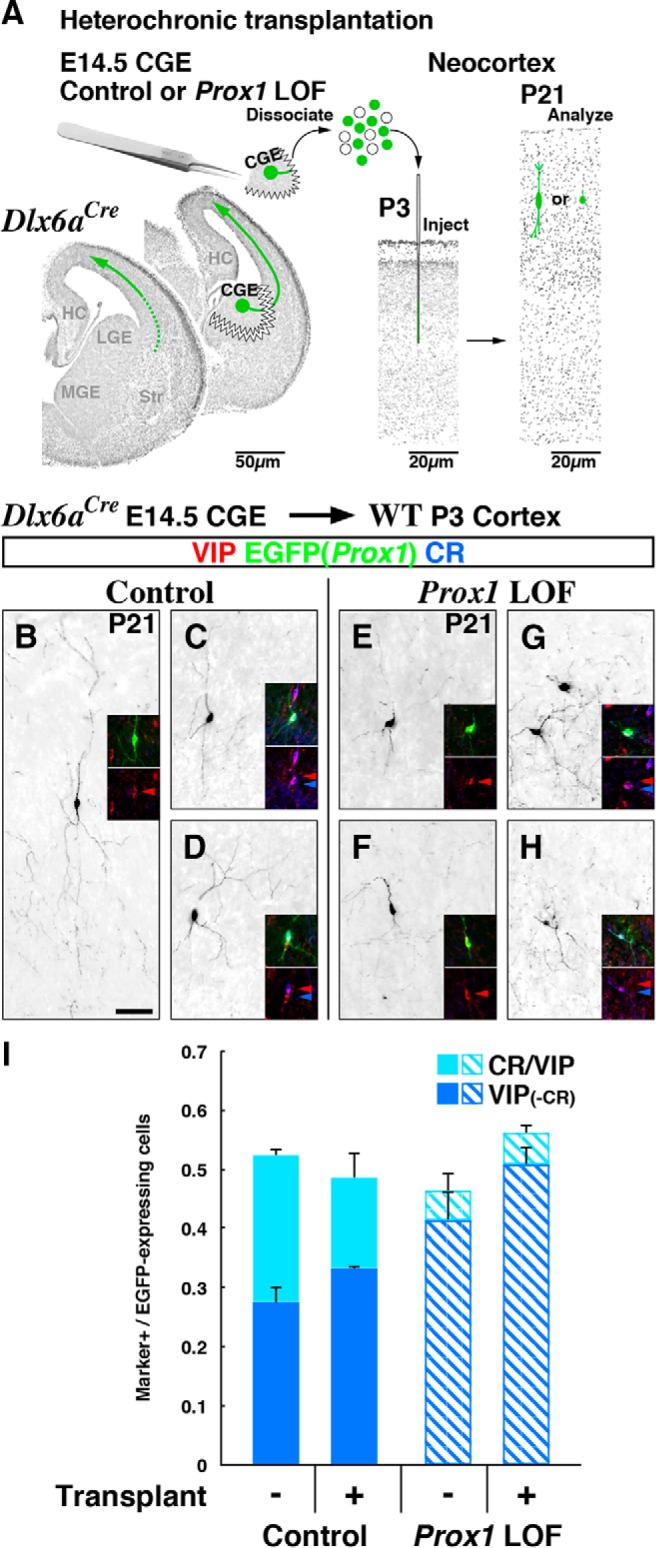
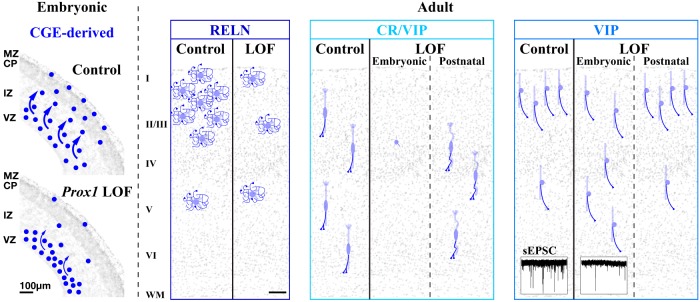
Neurogliaform (RELN+) and bipolar (VIP+) GABAergic interneurons of the mammalian cerebral cortex provide critical inhibition locally within the superficial layers. While these subtypes are known to originate from the embryonic caudal ganglionic eminence (CGE), the specific genetic programs that direct their positioning, maturation, and integration into the cortical network have not been elucidated. Here, we report that in mice expression of the transcription factor Prox1 is selectively maintained in postmitotic CGE-derived cortical interneuron precursors and that loss of Prox1 impairs the integration of these cells into superficial layers. Moreover, Prox1 differentially regulates the postnatal maturation of each specific subtype originating from the CGE (RELN, Calb2/VIP, and VIP). Interestingly, Prox1 promotes the maturation of CGE-derived interneuron subtypes through intrinsic differentiation programs that operate in tandem with extrinsically driven neuronal activity-dependent pathways. Thus Prox1 represents the first identified transcription factor specifically required for the embryonic and postnatal acquisition of CGE-derived cortical interneuron properties.
Significance statement: Despite the recognition that 30% of GABAergic cortical interneurons originate from the caudal ganglionic eminence (CGE), to date, a specific transcriptional program that selectively regulates the development of these populations has not yet been identified. Moreover, while CGE-derived interneurons display unique patterns of tangential and radial migration and preferentially populate the superficial layers of the cortex, identification of a molecular program that controls these events is lacking.Here, we demonstrate that the homeodomain transcription factor Prox1 is expressed in postmitotic CGE-derived cortical interneuron precursors and is maintained into adulthood. We found that Prox1 function is differentially required during both embryonic and postnatal stages of development to direct the migration, differentiation, circuit integration, and maintenance programs within distinct subtypes of CGE-derived interneurons.
Keywords: RELN; VIP; bipolar; mouse genetics; neurogliaform; transcription.
Copyright © 2015 the authors 0270-6474/15/3512870-21$15.00/0.
Figures
Similar articles
-
PROX1: a lineage tracer for cortical interneurons originating in the lateral/caudal ganglionic eminence and preoptic area.PLoS One. 2013 Oct 14;8(10):e77339. doi: 10.1371/journal.pone.0077339. eCollection 2013. PLoS One. 2013. PMID: 24155945 Free PMC article.
-
Genetic fate mapping reveals that the caudal ganglionic eminence produces a large and diverse population of superficial cortical interneurons.J Neurosci. 2010 Feb 3;30(5):1582-94. doi: 10.1523/JNEUROSCI.4515-09.2010. J Neurosci. 2010. PMID: 20130169 Free PMC article.
-
Neuronal activity is required for the development of specific cortical interneuron subtypes.Nature. 2011 Apr 21;472(7343):351-5. doi: 10.1038/nature09865. Epub 2011 Apr 3. Nature. 2011. PMID: 21460837 Free PMC article.
-
Transcriptional Regulation of Cortical Interneuron Development.In: Noebels JL, Avoli M, Rogawski MA, Vezzani A, Delgado-Escueta AV, editors. Jasper's Basic Mechanisms of the Epilepsies. 5th edition. New York: Oxford University Press; 2024. Chapter 47. In: Noebels JL, Avoli M, Rogawski MA, Vezzani A, Delgado-Escueta AV, editors. Jasper's Basic Mechanisms of the Epilepsies. 5th edition. New York: Oxford University Press; 2024. Chapter 47. PMID: 39637134 Free Books & Documents. Review.
-
Elucidating the developmental trajectories of GABAergic cortical interneuron subtypes.Neurosci Res. 2019 Jan;138:26-32. doi: 10.1016/j.neures.2018.09.012. Epub 2018 Sep 15. Neurosci Res. 2019. PMID: 30227162 Review.
Cited by
-
Role of DNMTs in the Brain.Adv Exp Med Biol. 2022;1389:363-394. doi: 10.1007/978-3-031-11454-0_15. Adv Exp Med Biol. 2022. PMID: 36350518 Review.
-
Developmental defects and impaired network excitability in a cerebral organoid model of KCNJ11 p.V59M-related neonatal diabetes.Sci Rep. 2021 Nov 3;11(1):21590. doi: 10.1038/s41598-021-00939-7. Sci Rep. 2021. PMID: 34732776 Free PMC article.
-
Identification of diverse astrocyte populations and their malignant analogs.Nat Neurosci. 2017 Mar;20(3):396-405. doi: 10.1038/nn.4493. Epub 2017 Feb 6. Nat Neurosci. 2017. PMID: 28166219 Free PMC article.
-
Involvement of cortical fast-spiking parvalbumin-positive basket cells in epilepsy.Prog Brain Res. 2016;226:81-126. doi: 10.1016/bs.pbr.2016.04.012. Epub 2016 Jun 7. Prog Brain Res. 2016. PMID: 27323940 Free PMC article. Review.
-
Optimizing Nervous System-Specific Gene Targeting with Cre Driver Lines: Prevalence of Germline Recombination and Influencing Factors.Neuron. 2020 Apr 8;106(1):37-65.e5. doi: 10.1016/j.neuron.2020.01.008. Epub 2020 Feb 5. Neuron. 2020. PMID: 32027825 Free PMC article.
References
-
- Anderson SA, Marín O, Horn C, Jennings K, Rubenstein JL. Distinct cortical migrations from the medial and lateral ganglionic eminences. Development. 2001;128:353–363. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases