

Deleted in Breast Cancer 1 Suppresses B Cell Activation through RelB and Is Regulated by IKKα Phosphorylation
- PMID: 26378077
- PMCID: PMC4642440
- DOI: 10.4049/jimmunol.1500713
Deleted in Breast Cancer 1 Suppresses B Cell Activation through RelB and Is Regulated by IKKα Phosphorylation
Abstract
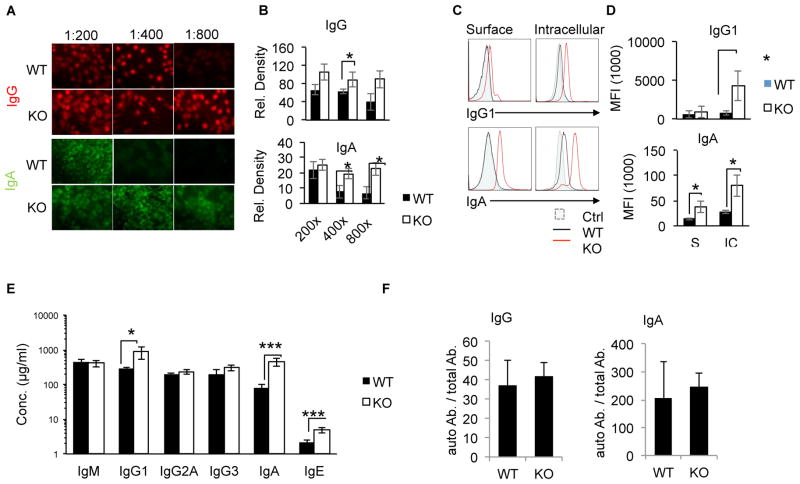
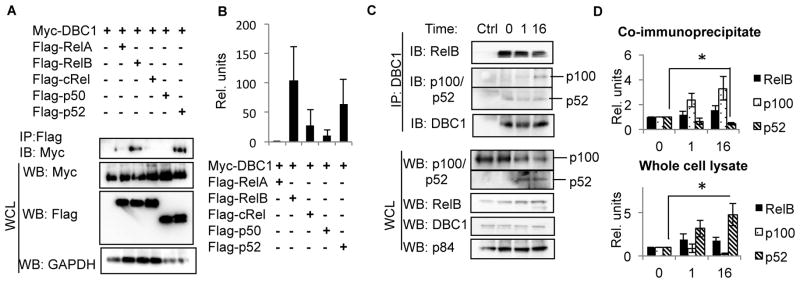
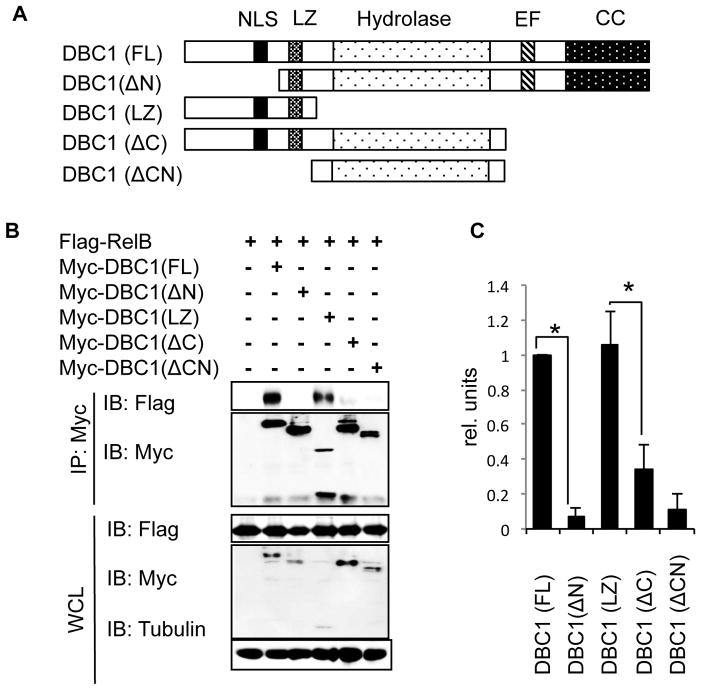
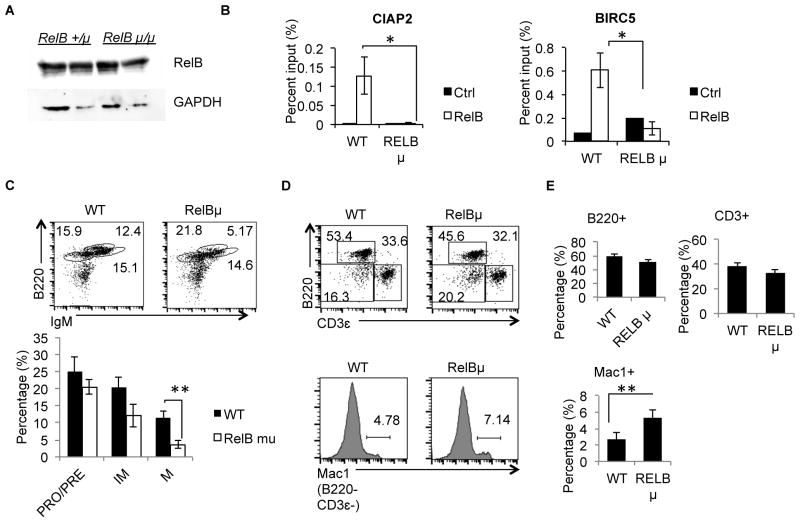
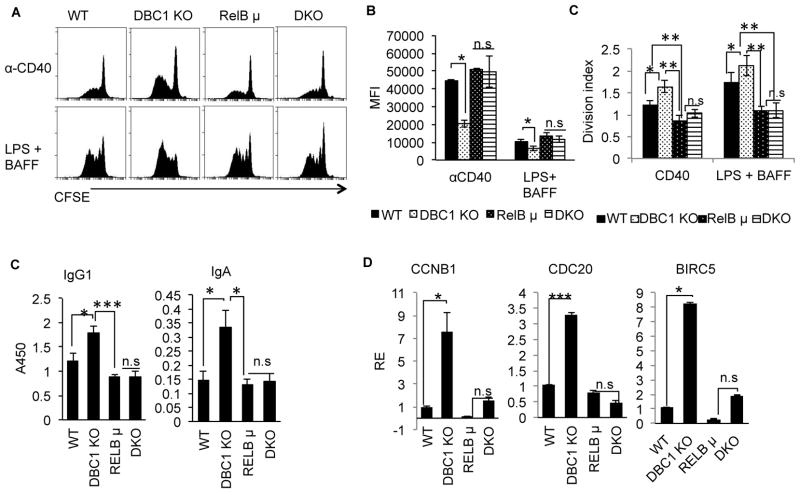
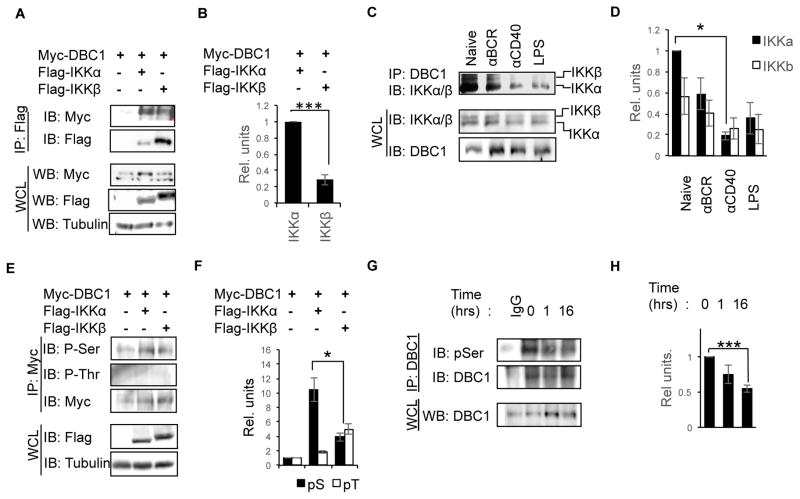
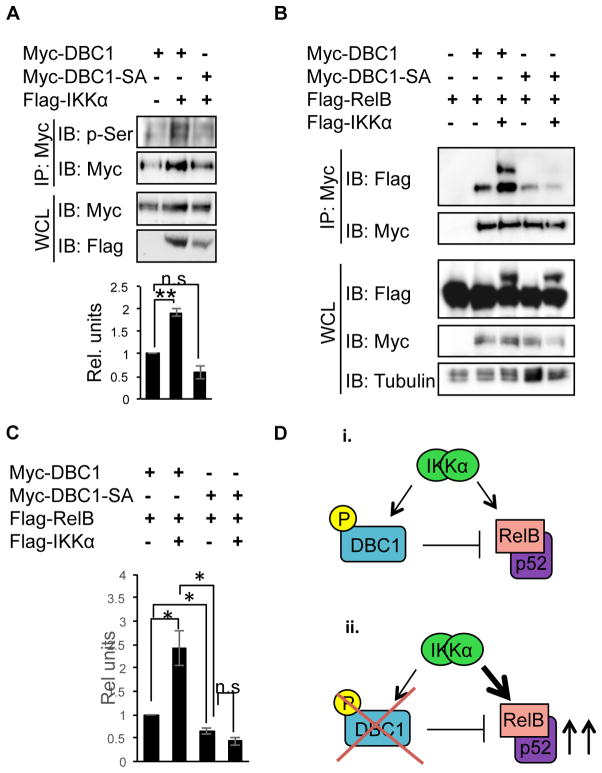
Alternative NF-κB signaling is crucial for B cell activation and Ig production, and it is mainly regulated by the inhibitor of κ B kinase (IKK) regulatory complex. Dysregulation of alternative NF-κB signaling in B cells could therefore lead to hyperactive B cells and Ig overproduction. In our previous, study we found that deleted in breast cancer 1 (DBC1) is a suppressor of the alternative NF-κB pathway to attenuate B cell activation. In this study, we report that loss of DBC1 results in spontaneous overproduction of Ig in mice after 10 mo of age. Using a double mutant genetic model, we confirm that DBC1 suppresses B cell activation through RelB inhibition. At the molecular level, we show that DBC1 interacts with alternative NF-κB members RelB and p52 through its leucine zipper domain. In addition, phosphorylation of DBC1 at its C terminus by IKKα facilitates its interaction with RelB and IKKα, indicating that DBC1-mediated suppression of alternative NF-κB is regulated by IKKα. Our results define the molecular mechanism of DBC1 inhibition of alternative NF-κB activation in suppressing B cell activation.
Copyright © 2015 by The American Association of Immunologists, Inc.
Conflict of interest statement
Disclosure of Conflicts of Interest. The authors declare no competing financial interests.
Figures
References
-
- Bonizzi G, Karin M. The two NF-[kappa]B activation pathways and their role in innate and adaptive immunity. Trends in Immunology. 2004;25:280–288. - PubMed
-
- Franzoso G, Carlson L, Poljak L, Shores EW, Epstein S, Leonardi A, Grinberg A, Tran T, Scharton-Kersten T, Anver M, Love P, Brown K, Siebenlist U. Mice Deficient in Nuclear Factor (NF)-κB/p52 Present with Defects in Humoral Responses, Germinal Center Reactions, and Splenic Microarchitecture. The Journal of Experimental Medicine. 1998;187:147–159. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
