

Receptor-like cytoplasmic kinase MARIS functions downstream of CrRLK1L-dependent signaling during tip growth
- PMID: 26378127
- PMCID: PMC4593096
- DOI: 10.1073/pnas.1512375112
Receptor-like cytoplasmic kinase MARIS functions downstream of CrRLK1L-dependent signaling during tip growth
Abstract
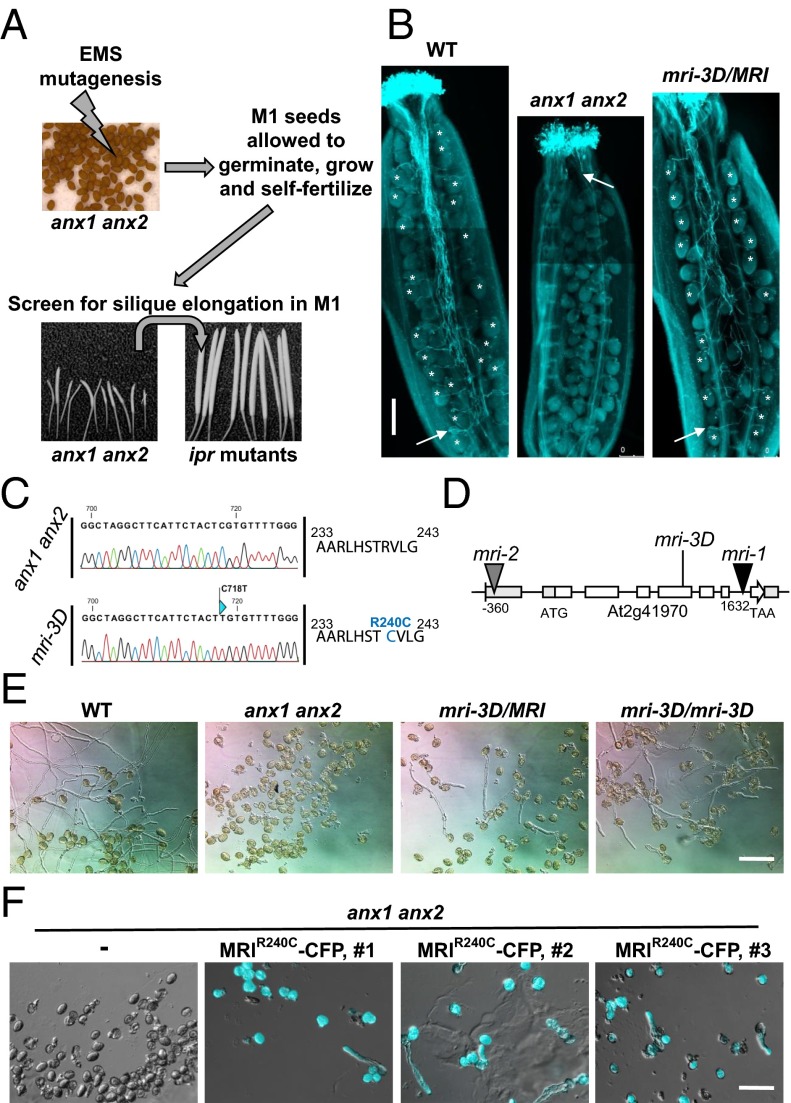
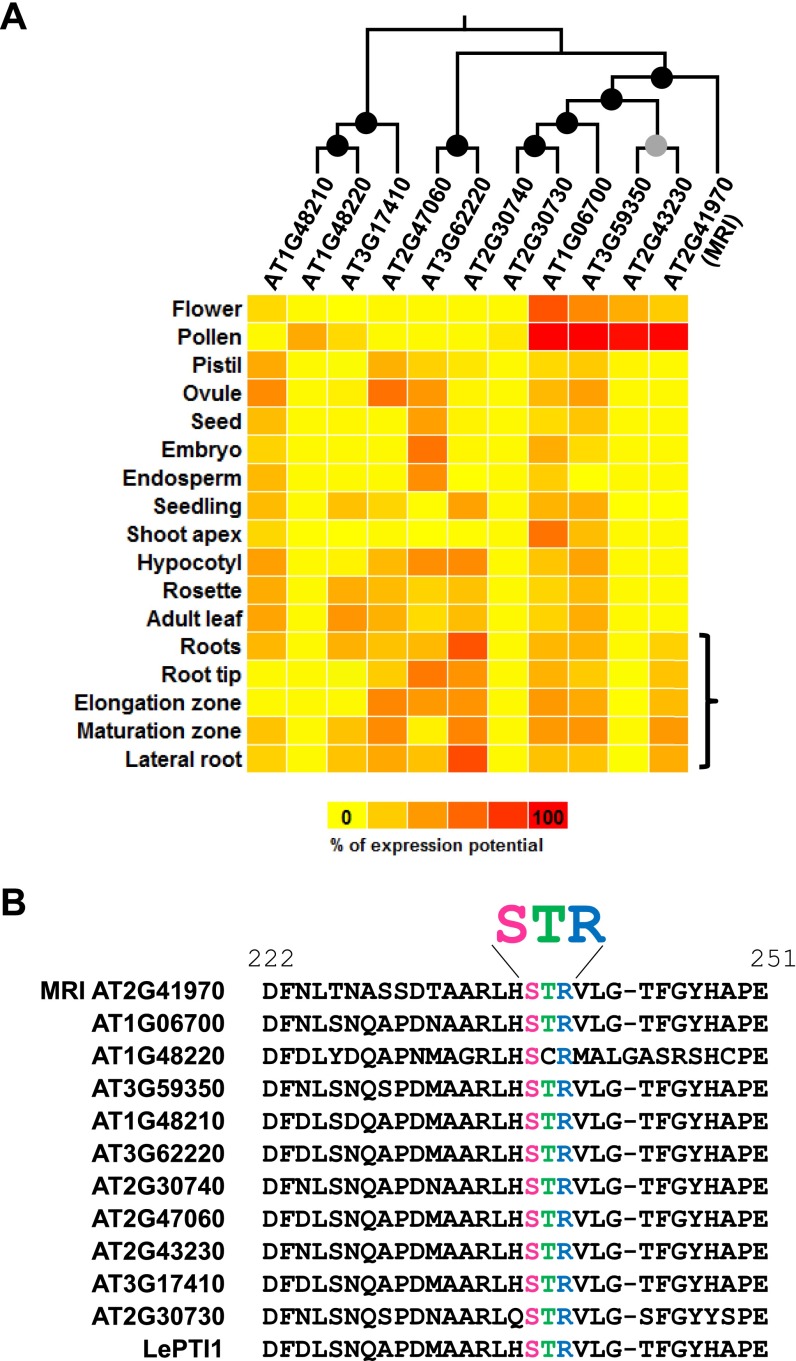
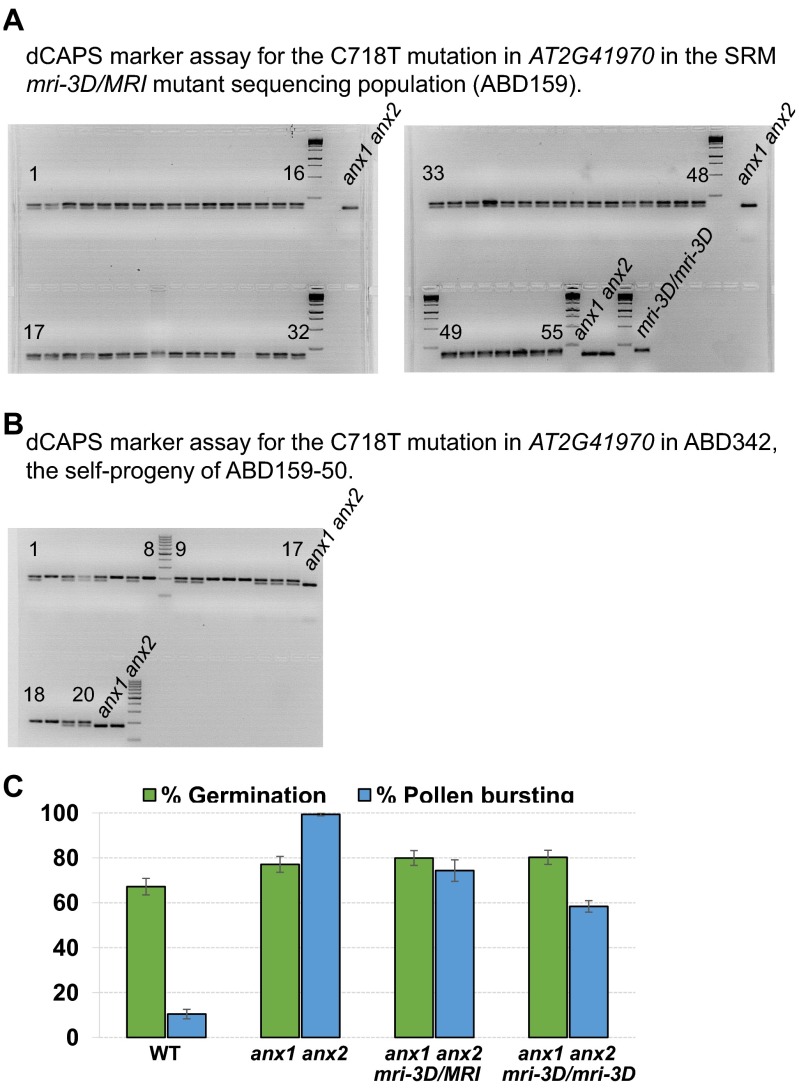
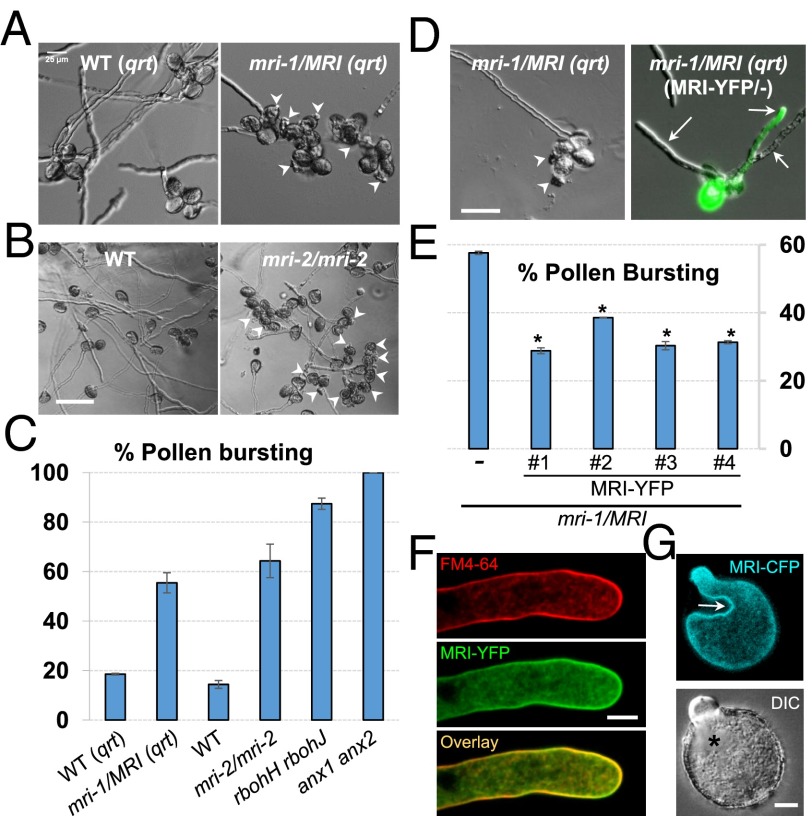
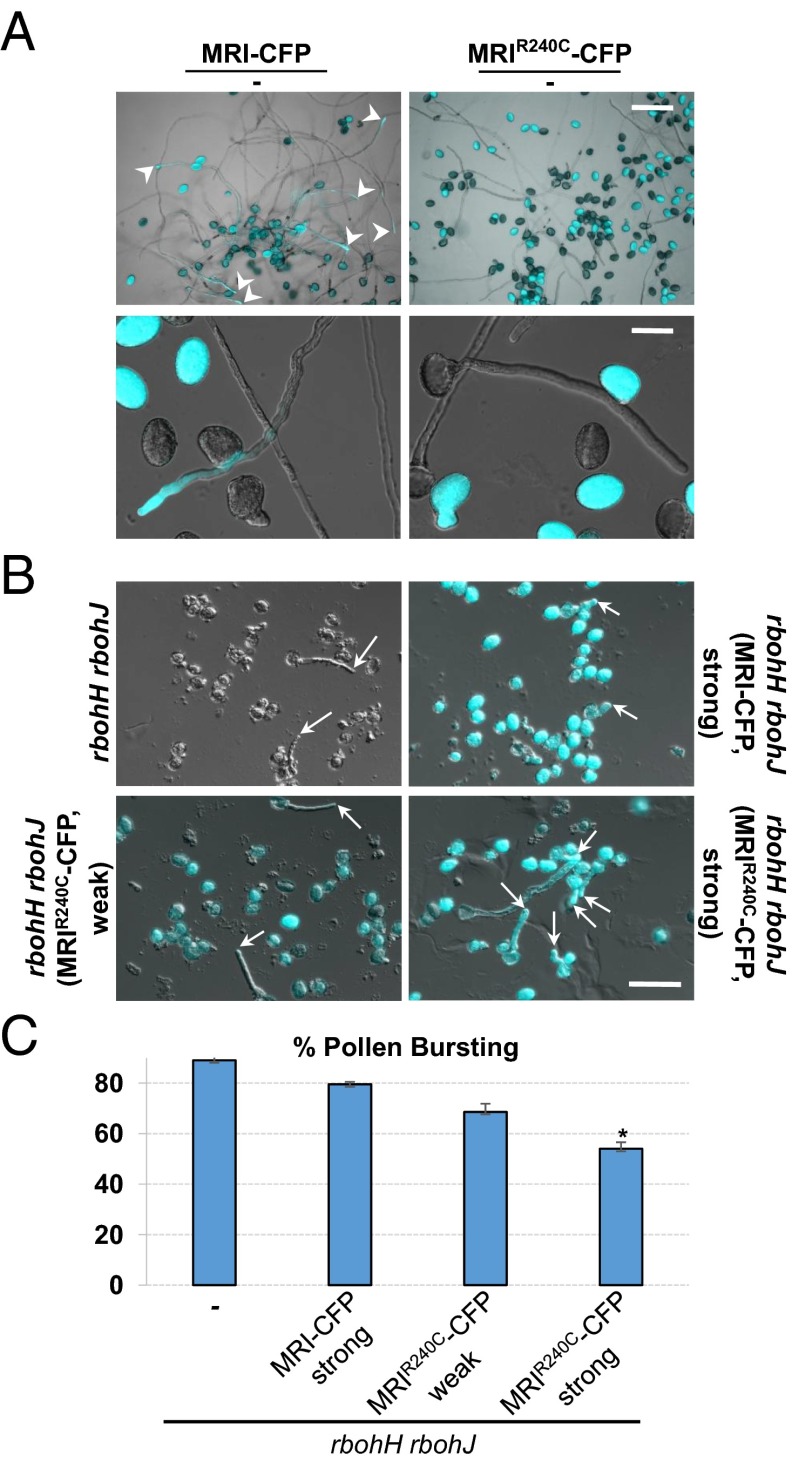
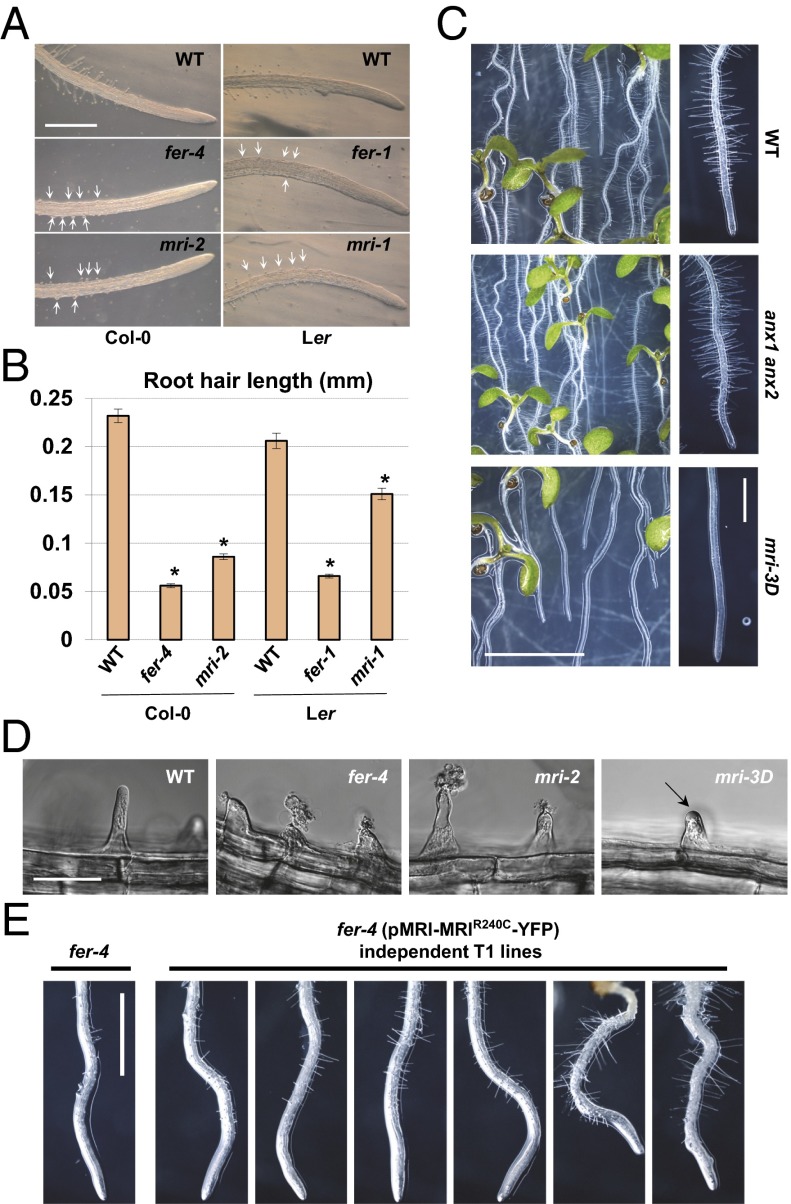
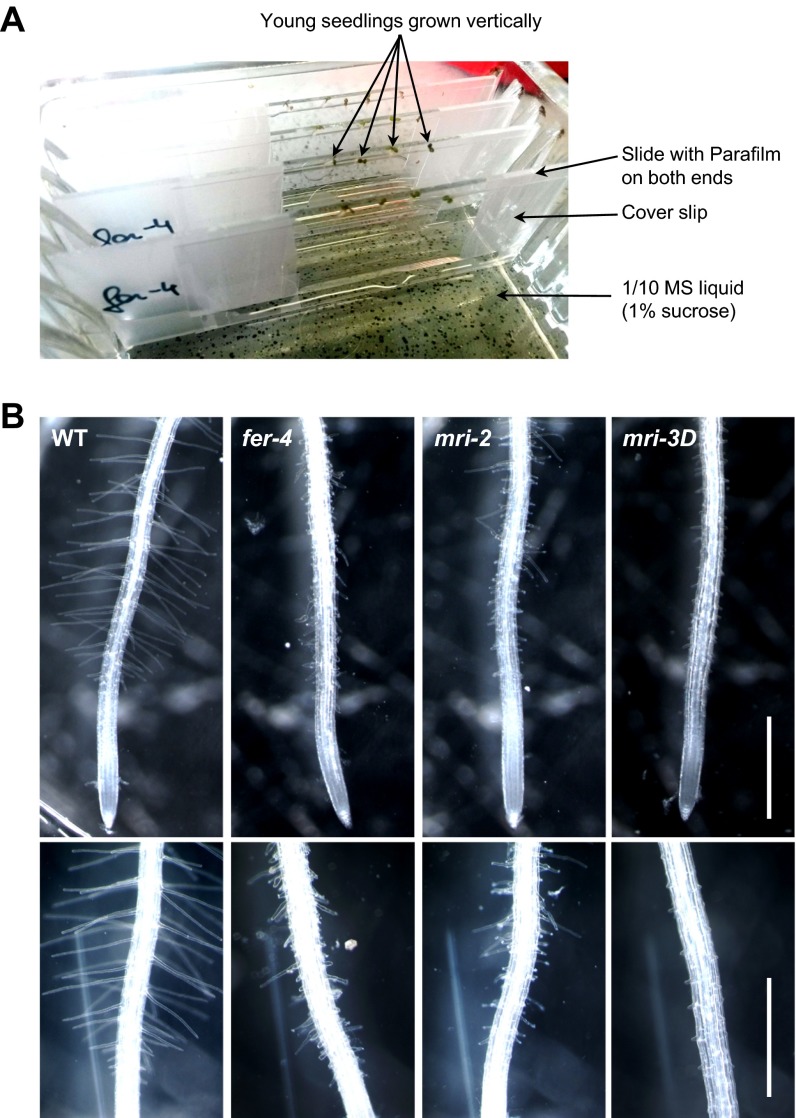
Growing plant cells need to rigorously coordinate external signals with internal processes. For instance, the maintenance of cell wall (CW) integrity requires the coordination of CW sensing with CW remodeling and biosynthesis to avoid growth arrest or integrity loss. Despite the involvement of receptor-like kinases (RLKs) of the Catharanthus roseus RLK1-like (CrRLK1L) subfamily and the reactive oxygen species-producing NADPH oxidases, it remains largely unknown how this coordination is achieved. ANXUR1 (ANX1) and ANX2, two redundant members of the CrRLK1L subfamily, are required for tip growth of the pollen tube (PT), and their closest homolog, FERONIA, controls root-hair tip growth. Previously, we showed that ANX1 overexpression mildly inhibits PT growth by oversecretion of CW material, whereas pollen tubes of anx1 anx2 double mutants burst spontaneously after germination. Here, we report the identification of suppressor mutants with improved fertility caused by the rescue of anx1 anx2 pollen tube bursting. Mapping of one these mutants revealed an R240C nonsynonymous substitution in the activation loop of a receptor-like cytoplasmic kinase (RLCK), which we named MARIS (MRI). We show that MRI is a plasma membrane-localized member of the RLCK-VIII subfamily and is preferentially expressed in both PTs and root hairs. Interestingly, mri-knockout mutants display spontaneous PT and root-hair bursting. Moreover, expression of the MRI(R240C) mutant, but not its wild-type form, partially rescues the bursting phenotypes of anx1 anx2 PTs and fer root hairs but strongly inhibits wild-type tip growth. Thus, our findings identify a novel positive component of the CrRLK1L-dependent signaling cascade that coordinates CW integrity and tip growth.
Keywords: Arabidopsis; cell wall integrity; pollen tube; receptor-like kinase signaling; root hair.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
