

Nitrite and nitrate-dependent generation of anti-inflammatory fatty acid nitroalkenes
- PMID: 26385079
- PMCID: PMC4684780
- DOI: 10.1016/j.freeradbiomed.2015.07.149
Nitrite and nitrate-dependent generation of anti-inflammatory fatty acid nitroalkenes
Abstract
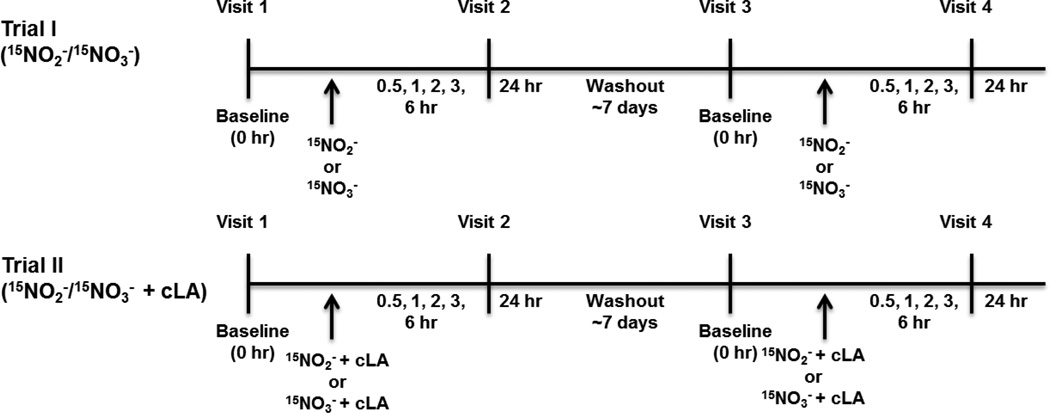
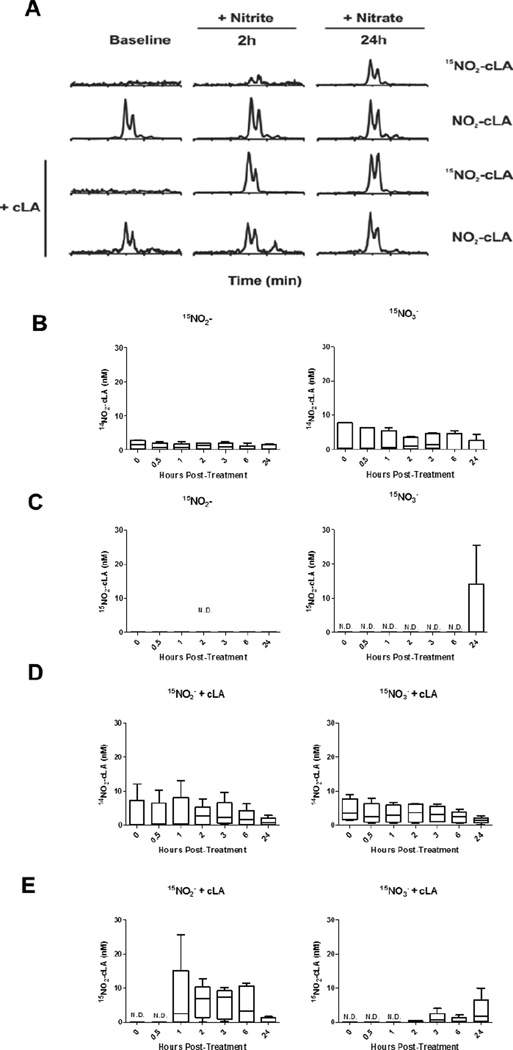
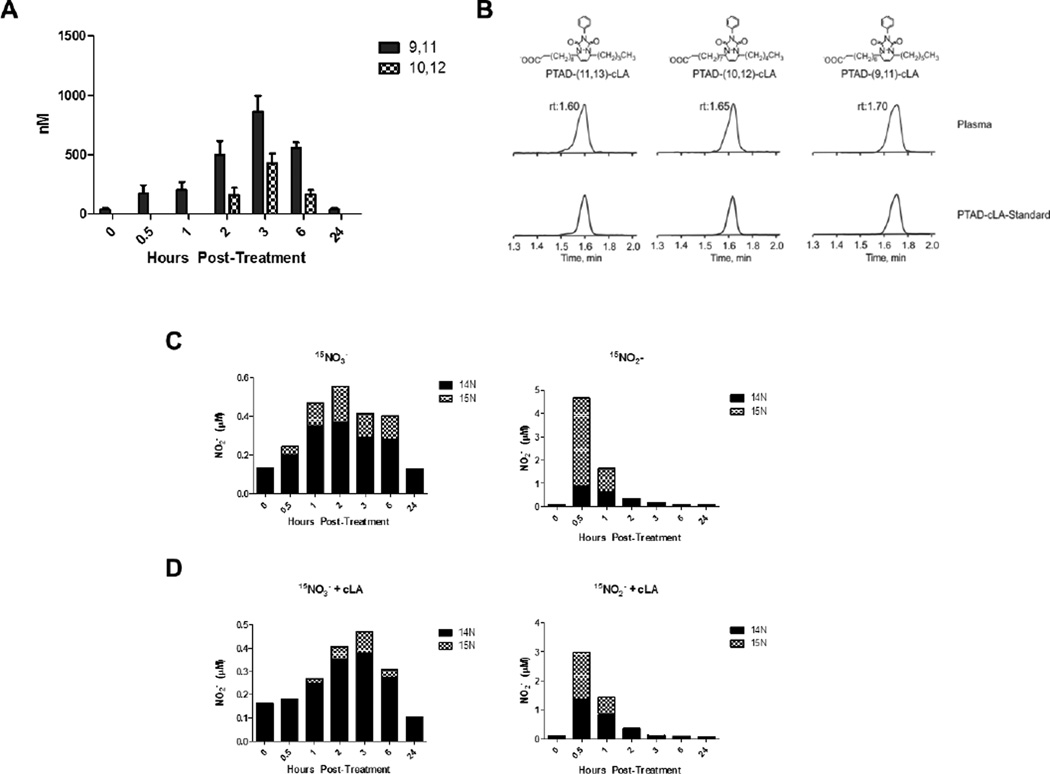
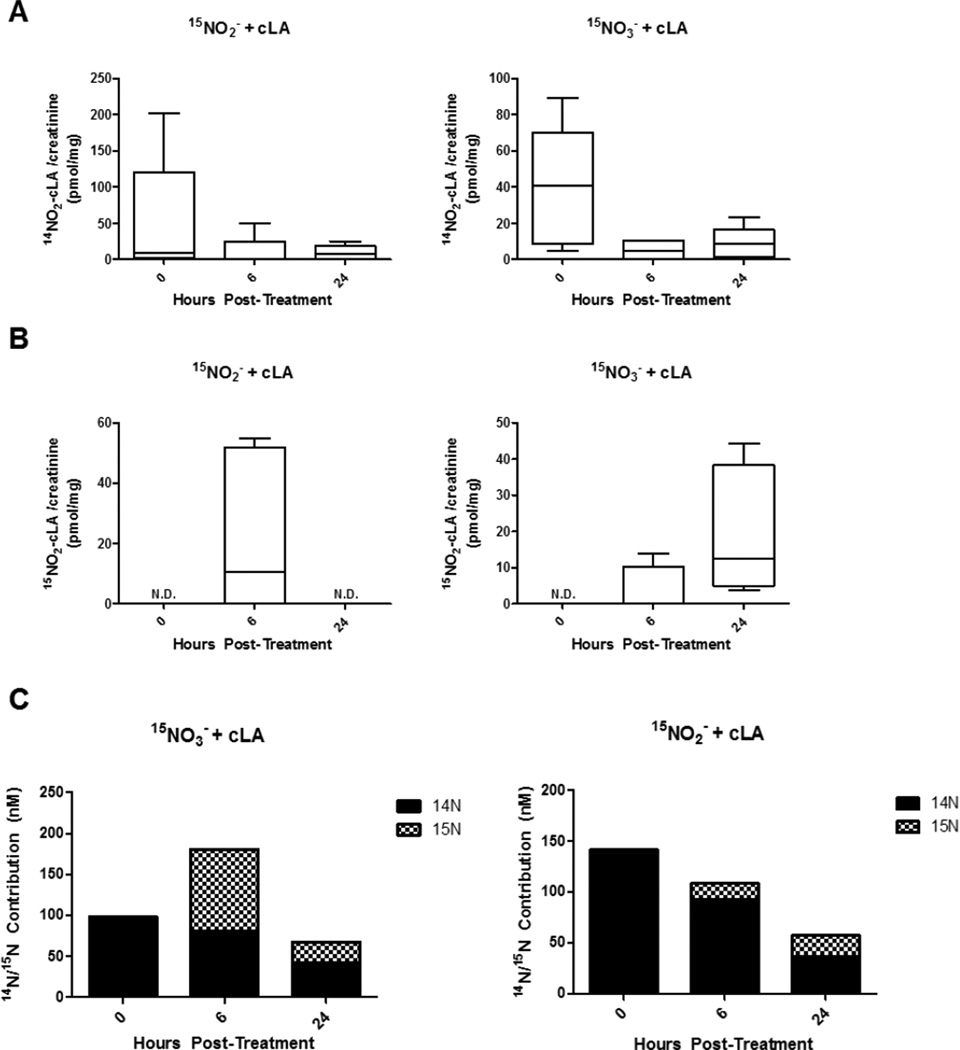
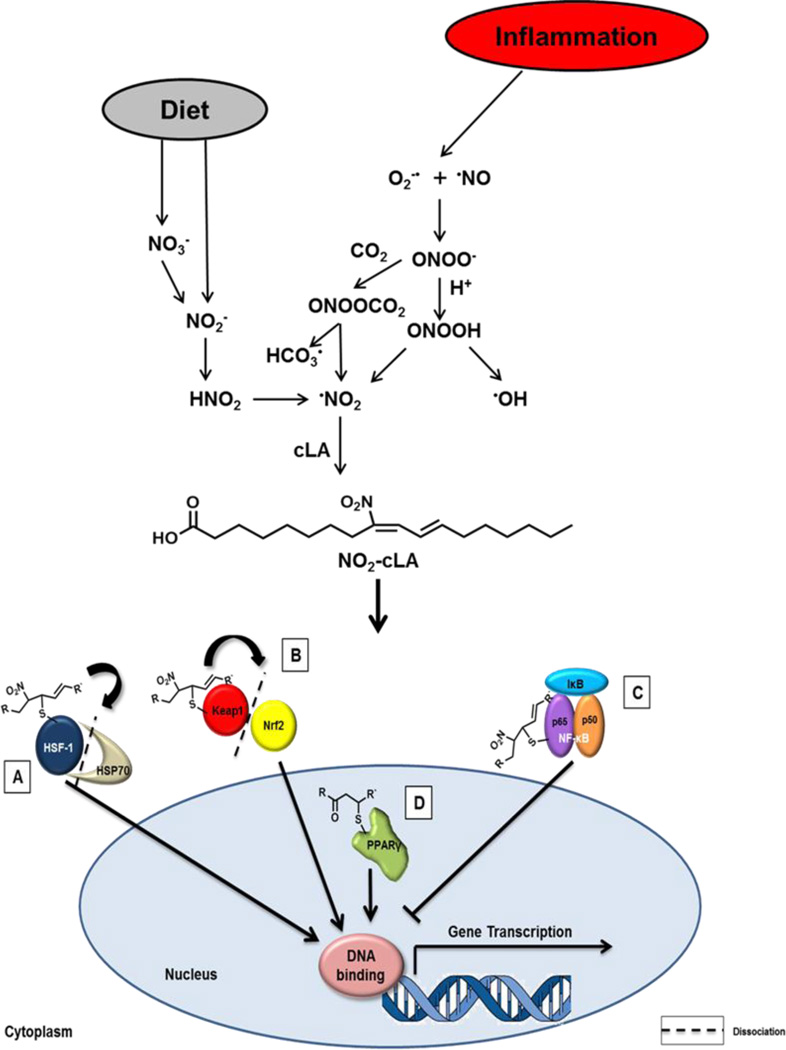
A gap in our understanding of the beneficial systemic responses to dietary constituents nitrate (NO3(-)), nitrite (NO2(-)) and conjugated linoleic acid (cLA) is the identification of the downstream metabolites that mediate their actions. To examine these reactions in a clinical context, investigational drug preparations of (15)N-labeled NO3(-) and NO2(-) were orally administered to healthy humans with and without cLA. Mass spectrometry analysis of plasma and urine indicated that the nitrating species nitrogen dioxide was formed and reacted with the olefinic carbons of unsaturated fatty acids to yield the electrophilic fatty acid, nitro-cLA (NO2-cLA). These species mediate the post-translational modification (PTM) of proteins via reversible Michael addition with nucleophilic amino acids. The PTM of critical target proteins by electrophilic lipids has been described as a sensing mechanism that regulates adaptive cellular responses, but little is known about the endogenous generation of fatty acid nitroalkenes and their metabolites. We report that healthy humans consuming (15)N-labeled NO3(-) or NO2(-), with and without cLA supplementation, produce (15)NO2-cLA and corresponding metabolites that are detected in plasma and urine. These data support that the dietary constituents NO3(-), NO2(-) and cLA promote the further generation of secondary electrophilic lipid products that are absorbed into the circulation at concentrations sufficient to exert systemic effects before being catabolized or excreted.
Keywords: Conjugated linoleic acid; Diet; Nitrate; Nitrite; Nitro-fatty acid; Nitrogen metabolism; Redox signaling.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
References
-
- Piskorski R, Hanus R, Vasickova S, Cvacka J, Sobotnik J, Svatos A, Valterova I. Nitroalkenes and sesquiterpene hydrocarbons from the frontal gland of three prorhinotermes termite species. Journal of chemical ecology. 2007;33:1787–1794. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 HL132550/HL/NHLBI NIH HHS/United States
- AT00682/AT/NCCIH NIH HHS/United States
- K12 HD052892/HD/NICHD NIH HHS/United States
- R25 AT000682/AT/NCCIH NIH HHS/United States
- R01 HL064937/HL/NHLBI NIH HHS/United States
- R01-HL64937/HL/NHLBI NIH HHS/United States
- P01-HL103455/HL/NHLBI NIH HHS/United States
- T32 HL110849/HL/NHLBI NIH HHS/United States
- UL1 TR000005/TR/NCATS NIH HHS/United States
- K23 HL124051/HL/NHLBI NIH HHS/United States
- P01 HL103455/HL/NHLBI NIH HHS/United States
- R37 HL058115/HL/NHLBI NIH HHS/United States
- R01-HL058115/HL/NHLBI NIH HHS/United States
- K12-HD052892/HD/NICHD NIH HHS/United States
- R01 HL058115/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
