

Biochemical analysis of hatchet self-cleaving ribozymes
- PMID: 26385510
- PMCID: PMC4604424
- DOI: 10.1261/rna.052522.115
Biochemical analysis of hatchet self-cleaving ribozymes
Abstract
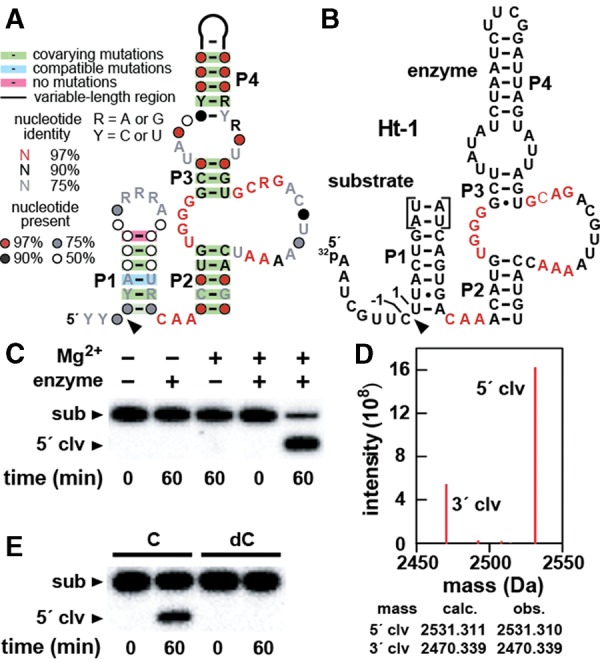
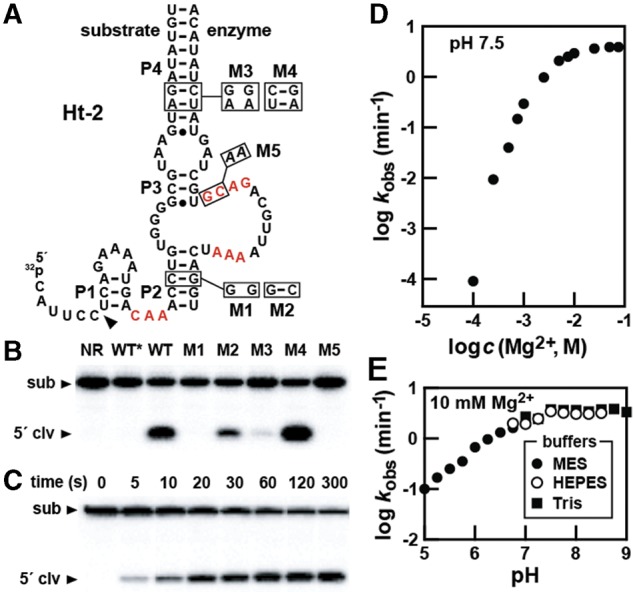
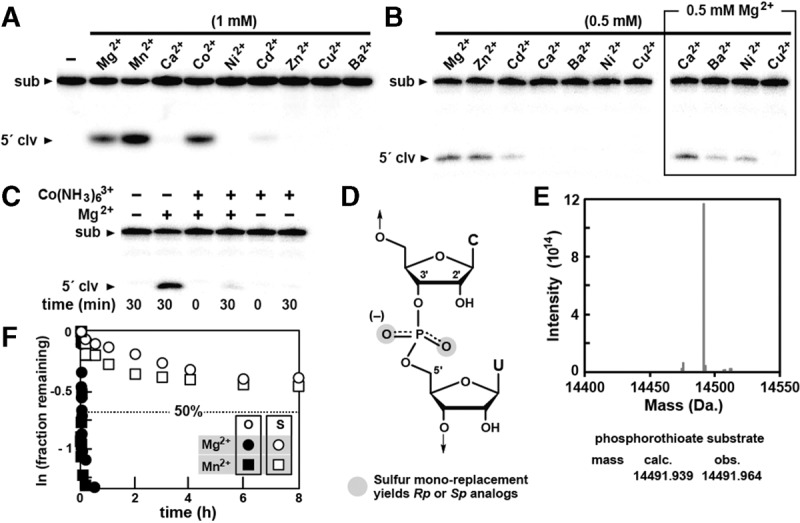
Hatchet RNAs are members of a novel self-cleaving ribozyme class that was recently discovered by using a bioinformatics search strategy. The consensus sequence and secondary structure of this class includes 13 highly conserved and numerous other modestly conserved nucleotides interspersed among bulges linking four base-paired substructures. A representative hatchet ribozyme from a metagenomic source requires divalent ions such as Mg(2+) to promote RNA strand scission with a maximum rate constant of ∼4 min(-1). As with all other small self-cleaving ribozymes discovered to date, hatchet ribozymes employ a general mechanism for catalysis involving the nucleophilic attack of a ribose 2'-oxygen atom on an adjacent phosphorus center. Kinetic characteristics of the reaction demonstrate that members of this ribozyme class have an essential requirement for divalent metal ions and that they might have a complex active site that employs multiple catalytic strategies to accelerate RNA cleavage by internal phosphoester transfer.
Keywords: RNA cleavage; RNA processing; comparative sequence analysis; phosphoester transfer; phosphorothioate.
© 2015 Li et al.; Published by Cold Spring Harbor Laboratory Press for the RNA Society.
Figures
Similar articles
-
Biochemical analysis of pistol self-cleaving ribozymes.RNA. 2015 Nov;21(11):1852-8. doi: 10.1261/rna.052514.115. Epub 2015 Sep 18. RNA. 2015. PMID: 26385507 Free PMC article.
-
Substrate specificity and reaction kinetics of an X-motif ribozyme.RNA. 2003 Jun;9(6):688-97. doi: 10.1261/rna.2600503. RNA. 2003. PMID: 12756327 Free PMC article.
-
Structural and Biochemical Properties of Novel Self-Cleaving Ribozymes.Molecules. 2017 Apr 24;22(4):678. doi: 10.3390/molecules22040678. Molecules. 2017. PMID: 28441772 Free PMC article. Review.
-
Biochemical analysis of cleavage and ligation activities of the pistol ribozyme from Paenibacillus polymyxa.RNA Biol. 2021 Nov;18(11):1858-1866. doi: 10.1080/15476286.2021.1874706. Epub 2021 Feb 23. RNA Biol. 2021. PMID: 33622172 Free PMC article.
-
Catalytic strategies of self-cleaving ribozymes.Acc Chem Res. 2008 Aug;41(8):1027-35. doi: 10.1021/ar800050c. Epub 2008 Jul 25. Acc Chem Res. 2008. PMID: 18652494 Review.
Cited by
-
Emergence of a "Cyclosome" in a Primitive Network Capable of Building "Infinite" Proteins.Life (Basel). 2019 Jun 18;9(2):51. doi: 10.3390/life9020051. Life (Basel). 2019. PMID: 31216720 Free PMC article.
-
Minimal twister sister-like self-cleaving ribozymes in the human genome revealed by deep mutational scanning.Elife. 2024 Dec 5;12:RP90254. doi: 10.7554/eLife.90254. Elife. 2024. PMID: 39636683 Free PMC article.
-
Thirty-five years of research into ribozymes and nucleic acid catalysis: where do we stand today?F1000Res. 2016 Jun 27;5:F1000 Faculty Rev-1511. doi: 10.12688/f1000research.8601.1. eCollection 2016. F1000Res. 2016. PMID: 27408700 Free PMC article. Review.
-
Structure-based mechanistic insights into catalysis by small self-cleaving ribozymes.Curr Opin Chem Biol. 2017 Dec;41:71-83. doi: 10.1016/j.cbpa.2017.09.017. Epub 2017 Nov 3. Curr Opin Chem Biol. 2017. PMID: 29107885 Free PMC article. Review.
-
Novel ribozymes: discovery, catalytic mechanisms, and the quest to understand biological function.Nucleic Acids Res. 2019 Oct 10;47(18):9480-9494. doi: 10.1093/nar/gkz737. Nucleic Acids Res. 2019. PMID: 31504786 Free PMC article.
References
-
- Canny MD, Jucker FM, Kellogg E, Khvorova A, Jayasena SD, Pardi A. 2004. Fast cleavage kinetics of a natural hammerhead ribozyme. J Am Chem Soc 126: 10848–10849. - PubMed
-
- Cowan JA. 1993. Metallobiochemistry of RNA. Co(NH3)63+ as a probe for Mg2+(aq) binding sites. J Inorg Biochem 49: 171–175. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous