

Functional constraints in the evolution of brain circuits
- PMID: 26388716
- PMCID: PMC4555059
- DOI: 10.3389/fnins.2015.00303
Functional constraints in the evolution of brain circuits
Abstract
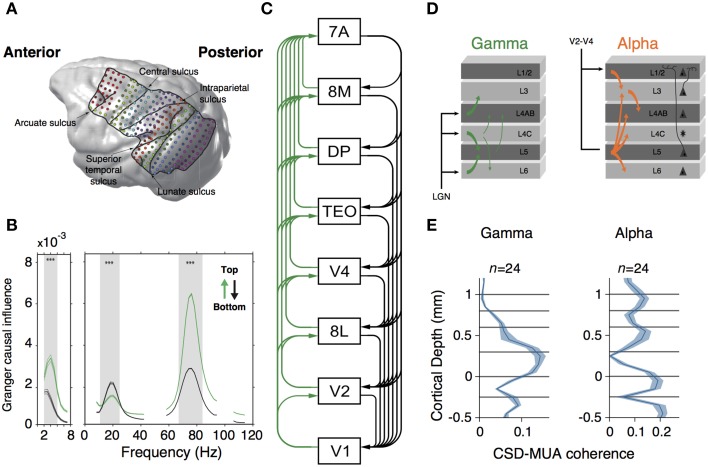
Regardless of major anatomical and neurodevelopmental differences, the vertebrate isocortex shows a remarkably well-conserved organization. In the isocortex, reciprocal connections between excitatory and inhibitory neurons are distributed across multiple layers, encompassing modular, dynamical and recurrent functional networks during information processing. These dynamical brain networks are often organized in neuronal assemblies interacting through rhythmic phase relationships. Accordingly, these oscillatory interactions are observed across multiple brain scale levels, and they are associated with several sensory, motor, and cognitive processes. Most notably, oscillatory interactions are also found in the complete spectrum of vertebrates. Yet, it is unknown why this functional organization is so well conserved in evolution. In this perspective, we propose some ideas about how functional requirements of the isocortex can account for the evolutionary stability observed in microcircuits across vertebrates. We argue that isocortex architectures represent canonical microcircuits resulting from: (i) the early selection of neuronal architectures based on the oscillatory excitatory-inhibitory balance, which lead to the implementation of compartmentalized oscillations and (ii) the subsequent emergence of inferential coding strategies (predictive coding), which are able to expand computational capacities. We also argue that these functional constraints may be the result of several advantages that oscillatory activity contributes to brain network processes, such as information transmission and code reliability. In this manner, similarities in mesoscale brain circuitry and input-output organization between different vertebrate groups may reflect evolutionary constraints imposed by these functional requirements, which may or may not be traceable to a common ancestor.
Keywords: canonical microcircuits; cortical evolution; cortical neurodevelopment; neuronal oscillations; predictive coding.
Figures


References
-
- Aboitiz F. (1995). Homology in the evolution of the cerebral hemispheres. The case of reptilian dorsal ventricular ridge and its possible correspondence with mammalian neocortex. J. Hirnforsch. 36, 461–472. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources