

Review
doi: 10.3389/fncel.2015.00340.
eCollection 2015.
Oligodendrocytes in a Nutshell
Affiliations
- PMID: 26388730
- PMCID: PMC4556025
- DOI: 10.3389/fncel.2015.00340
Item in Clipboard
Review
Oligodendrocytes in a Nutshell
Front Cell Neurosci.
.
Abstract
Oligodendrocytes are the myelinating cells of the central nervous system (CNS). While the phrase is oft repeated and holds true, the last few years have borne witness to radical change in our understanding of this unique cell type. Once considered static glue, oligodendrocytes are now seen as plastic and adaptive, capable of reacting to a changing CNS. This review is intended as a primer and guide, exploring how the past 5 years have fundamentally altered our appreciation of oligodendrocyte development and CNS myelination.
Keywords: OPC; actin; cytoskeleton; glial cell; microtubule; myelin; myelination.
Figures
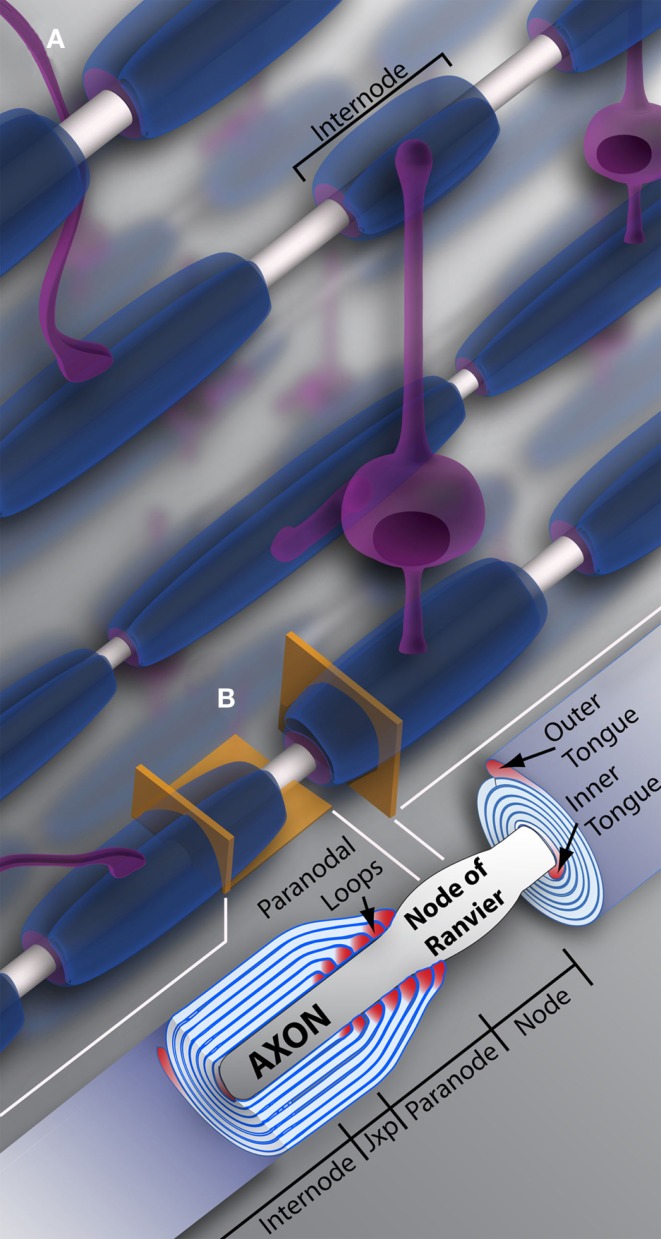
Oligodendrocytes and myelin. (A) Schematic representation of oligodendrocytes and myelin internodes in the CNS. As depicted, a typical oligodendrocyte (in purple) is capable of myelinating multiple axons (depicted in light gray). The oligodendrocyte will extend numerous processes, each ending in a myelin internode, depicted here in blue. Internodes within the same region, even along the same axon, can differ in length and/or size, providing the neuronal network or individual neurons with unique myelin profiles. (B) Vertical and longitudinal sections through a myelin sheath. Note the cytoplasm rich inner and outer tongues. The axon is unmyelinated and bulges at the Node of Ranvier, also referred to as the node. Swollen cytoplasmic rich paranodal loops (cytoplasm here denoted by the color pink) abut the axon, delineating the paranode. Paranodal loops anchor the myelin membrane to the axolemma through axo-glial junctions. Junctions are composed of axolemma and OL-based proteins, notably contactin, contactin-associated protein (Caspr), and neurofascin 155. These proteins hook together, keeping the loops and axolemma in close proximity. The paranode gives way to the juxtaparanode (juxtaparanode, as it is in juxtaposition with the paranode), a small area rich in K+ channels. Next is the internode, that can refer both to the myelin sheath as a whole (A), or the space under compact myelin between juxtaparanodes. Jxp = juxtaparanode.
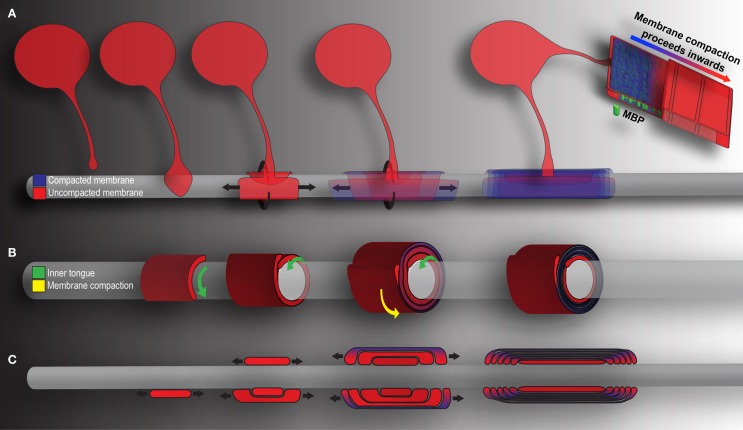
Mechanics of CNS myelination. Presented is a simplified schematic of OL-mediated myelination from various points of view. Cytoplasm rich, newly deposited membrane is represented in red, while compact membrane is shown in blue. (A) OLs extend a “growth-cone-like” tip seeking bare axons. Upon axonal contact, the tip expands, forming a triangular shaped membrane. Membrane growth occurs through simultaneous radial (around) and lateral (along) expansion. The growing inner tongue pushes radially under pre-existing membrane, depositing new membrane in its wake. During this process, the outermost layers of membrane (those first deposited) fill with MBP and compaction begins. Compaction and associated MBP clustering follows behind the growing inner tongue, zippering the newly formed membrane shut. When all necessary membrane has been deposited, the inner tongue halts growth and enters a period of stasis (shrinking as a result). It can be reactivated at a later time point to deposit additional myelin as required by the axon. (B) A cross-section through myelin membrane as presented in (A). Again, the inner tongue grows radially underneath pre-existing membrane. Membrane compaction occurs first in the older, outermost membrane layers and proceeds inwards. (C) Longitudinal representation of myelin membrane as presented in (A). Note that the lateral growth and eventual formation of paranodal loops as the outermost layers compact to form mature myelin.
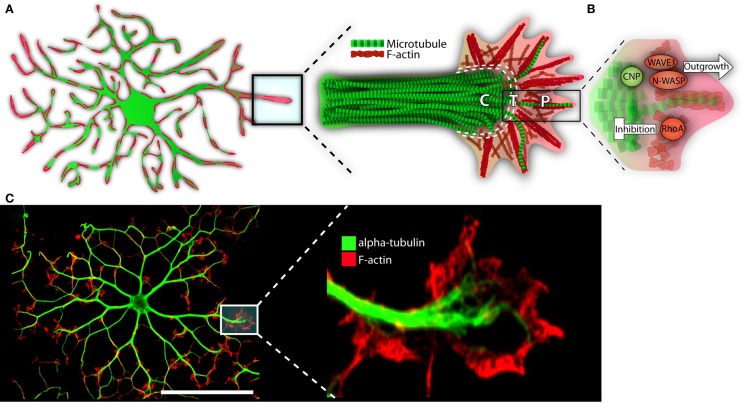
The growth-like cone and cytoskeleton in maturing oligodendrocytes. (A) Microtubules (depicted in green) run the length of OL processes in long parallel lines. F-actin (in red) is found throughout the OL, but is most heavily concentrated at the growing ends. These growing ends, or tips, are highly reminiscent of neuronal growth cones in both structure and cytoskeletal distribution. Neural cones have three distinct domains: a microtubule rich central (C) domain, an F-actin rich peripheral (P) domain, and a transient (T) domain where microtubules and F-actin overlap. We have overlaid these domains with the OL growth-cone-like structure to demonstrate similarities. In the schematic, microtubules (in green) invade the OL growth-cone centrally. Most do not advance much past this central point and are clearly delineated from the peripheral F-actin. The OL’s leading edge is replete with a meshwork of F-actin fibers (depicted in red). As well, parallel F-actin bundles (depicted as bundled long red strands), push against the membrane edge generating filopodia. A very few microtubules will penetrate the peripheral domain. In a neuronal cone, these exploratory microtubules are important for directed growth. (B) A basic schematic depicting various cytoskeletal assembly/remodeling proteins that drive or stall the oligodendrocyte’s growth cone. WAVE1 and N-WASP widen the cones lamellopodia, leading to process extension through F-actin branching and polymerization. CNP directs F-actin rearrangement, microtubule growth/bundling, as well as acts as a bridge between these two cytoskeletal components. CNP’s expression is strongly linked to OL morphogenesis; its ability to synchronize dynamic F-actin reorganization with microtubule polymerization and, therefore, process stabilization, is in large part responsible for OL outgrowth. RhoA, by contrast, acts as a stop-gate, preventing growth through generation of actomyosin contractile forces. Only when the RhoA pathway is deactivated can morphological development proceed. (C) Immunofluorescent representation of cytoskeleton and growth-like cone in the OL. The left panel depicts an immature OL with F-actin in red and α-tubulin in green. The right depicts a single growth-cone-like structure on the tip of an elongated OL process. P = peripheral, C = central, T = transient. Scale bar: 50 μm.
Similar articles
-
Evolutionary Origins of the Oligodendrocyte Cell Type and Adaptive Myelination.Front Neurosci. 2021 Dec 1;15:757360. doi: 10.3389/fnins.2021.757360. eCollection 2021. Front Neurosci. 2021. PMID: 34924932 Free PMC article. Review.
-
Oligodendrocytes in Development, Myelin Generation and Beyond.Cells. 2019 Nov 12;8(11):1424. doi: 10.3390/cells8111424. Cells. 2019. PMID: 31726662 Free PMC article. Review.
-
Cytoskeletal Linker Protein Dystonin Is Not Critical to Terminal Oligodendrocyte Differentiation or CNS Myelination.PLoS One. 2016 Feb 17;11(2):e0149201. doi: 10.1371/journal.pone.0149201. eCollection 2016. PLoS One. 2016. PMID: 26886550 Free PMC article.
-
In vivo actions of fibroblast growth factor-2 and insulin-like growth factor-I on oligodendrocyte development and myelination in the central nervous system.J Neurosci Res. 1999 Jul 1;57(1):74-85. doi: 10.1002/(SICI)1097-4547(19990701)57:1<74::AID-JNR8>3.0.CO;2-O. J Neurosci Res. 1999. PMID: 10397637
-
Neuronal activity and AMPA-type glutamate receptor activation regulates the morphological development of oligodendrocyte precursor cells.Glia. 2015 Jun;63(6):1021-35. doi: 10.1002/glia.22799. Epub 2015 Mar 4. Glia. 2015. PMID: 25739948
Cited by
-
Impact of neonatal hypoxia-ischaemia on oligodendrocyte survival, maturation and myelinating potential.J Cell Mol Med. 2018 Jan;22(1):207-222. doi: 10.1111/jcmm.13309. Epub 2017 Aug 7. J Cell Mol Med. 2018. PMID: 28782169 Free PMC article.
-
Role of glia in optic nerve.Prog Retin Eye Res. 2021 Mar;81:100886. doi: 10.1016/j.preteyeres.2020.100886. Epub 2020 Aug 6. Prog Retin Eye Res. 2021. PMID: 32771538 Free PMC article. Review.
-
Daam2 Regulates Myelin Structure and the Oligodendrocyte Actin Cytoskeleton through Rac1 and Gelsolin.J Neurosci. 2022 Mar 2;42(9):1679-1691. doi: 10.1523/JNEUROSCI.1517-21.2022. Epub 2022 Jan 31. J Neurosci. 2022. PMID: 35101966 Free PMC article.
-
Mechanistic Target of Rapamycin Regulates the Oligodendrocyte Cytoskeleton during Myelination.J Neurosci. 2020 Apr 8;40(15):2993-3007. doi: 10.1523/JNEUROSCI.1434-18.2020. Epub 2020 Mar 5. J Neurosci. 2020. PMID: 32139584 Free PMC article.
-
Oligodendrocytes in the aging brain.Neuronal Signal. 2021 Jul 6;5(3):NS20210008. doi: 10.1042/NS20210008. eCollection 2021 Sep. Neuronal Signal. 2021. PMID: 34290887 Free PMC article. Review.
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous