

KDM4B and KDM4A promote endometrial cancer progression by regulating androgen receptor, c-myc, and p27kip1
- PMID: 26397136
- PMCID: PMC4741634
- DOI: 10.18632/oncotarget.5165
KDM4B and KDM4A promote endometrial cancer progression by regulating androgen receptor, c-myc, and p27kip1
Abstract
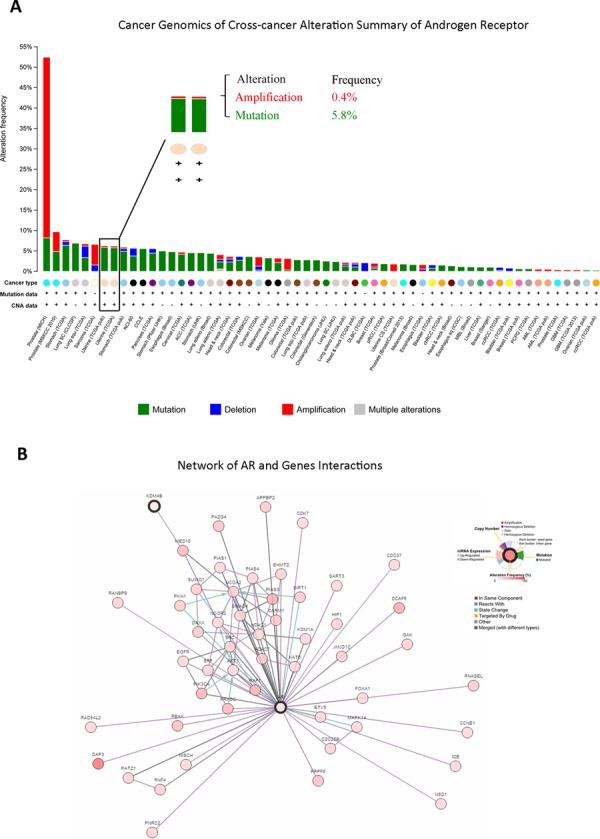
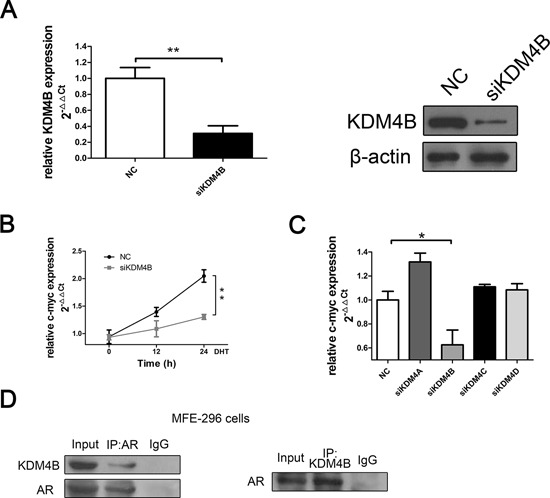
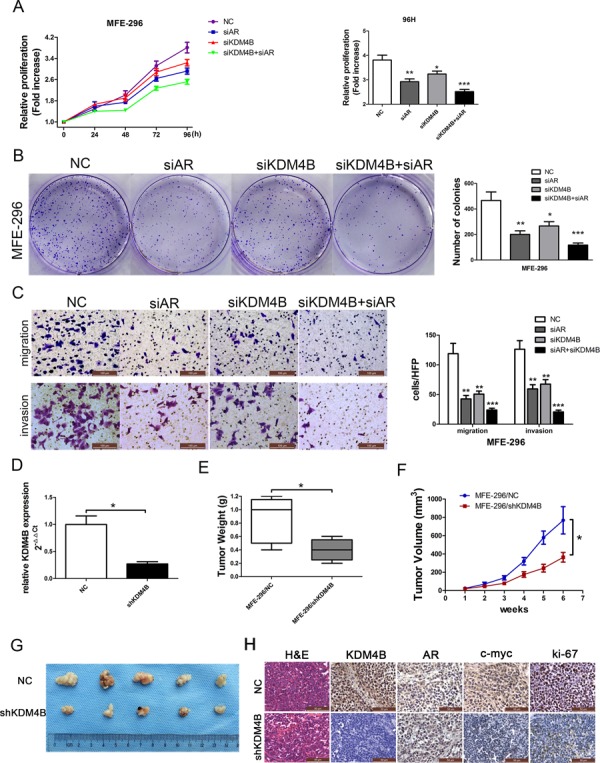
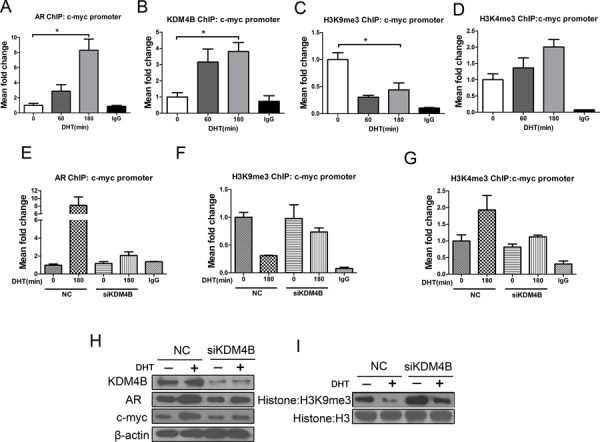
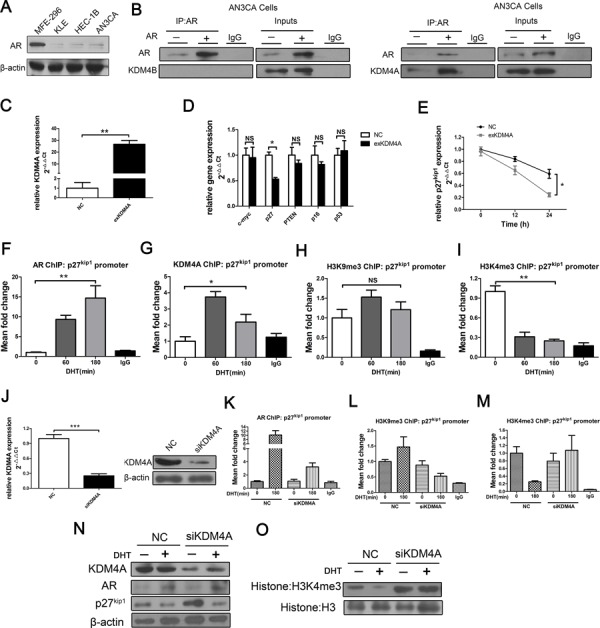
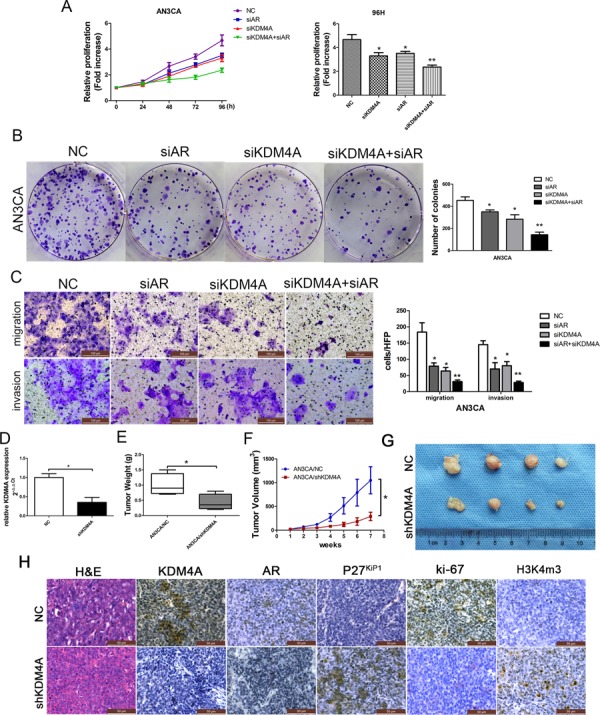
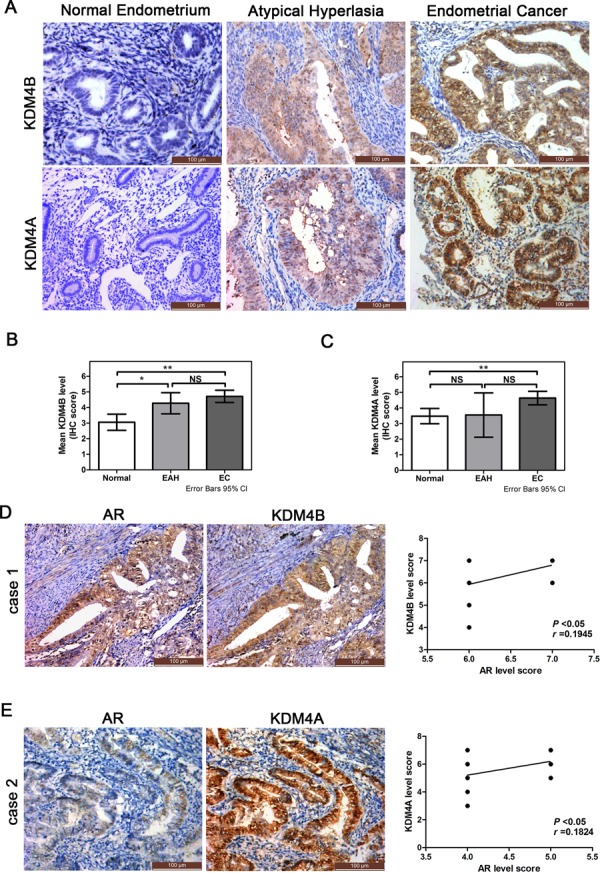
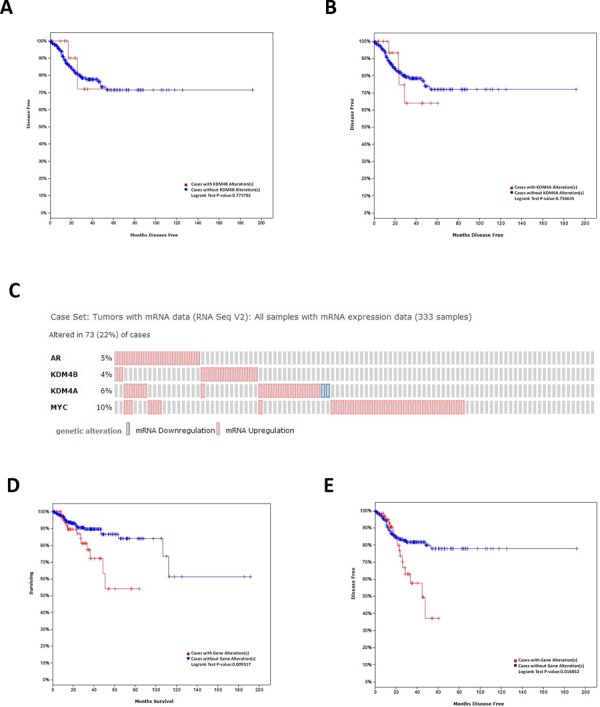
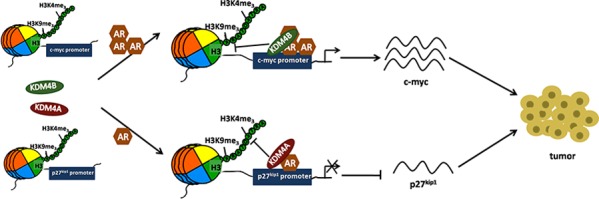
Epidemiological evidence suggests that elevated androgen levels and genetic variation related to the androgen receptor (AR) increase the risk of endometrial cancer (EC). However, the role of AR in EC is poorly understood. We report that two members of the histone demethylase KDM4 family act as major regulators of AR transcriptional activityin EC. In the MFE-296 cell line, KDM4B and AR upregulate c-myc expression, while in AN3CA cells KDM4A and AR downregulate p27kip1. Additionally, KDM4B expression is positively correlated with AR expression in EC cell lines with high baseline AR expression, while KDM4A and AR expression are positively correlated in low-AR cell lines. In clinical specimens, both KDM4B and KDM4A expression are significantly higher in EC tissues than that in normal endometrium. Finally, patients with alterations in AR, KDM4B, KDM4A, and c-myc have poor overall and disease-free survival rates. Together, these findings demonstrate that KDM4B and KDM4A promote EC progression by regulating AR activity.
Keywords: androgen receptor; c-myc; endometrial cancer; histone modification; lysine demethylases KDM4B and KDM4A; p27kip1; prognosis.
Conflict of interest statement
The authors declare that they have no conflicts of interest.
Figures
References
-
- Siegel RL, Miller KD, Jemal A. Cancer statistics, 2015. CA Cancer J Clin. 2015;65:5–29. - PubMed
-
- Whitney CW, Brunetto VL, Zaino RJ, Lentz SS, Sorosky J, Armstrong DK, Lee RB. Phase II study of medroxyprogesterone acetate plus tamoxifen in advanced endometrial carcinoma: a Gynecologic Oncology Group study. Gynecol Oncol. 2004;92:4–9. - PubMed
-
- Erkanli S, Ayhan A. Fertility-sparing therapy in young women with endometrial cancer: update. Int J Gynecol Cancer. 2010;20:1170–1187. - PubMed
-
- Fearnley EJ, Marquart L, Spurdle AB, Weinstein P, Webb PM. Polycystic ovary syndrome increases the risk of endometrial cancer in women aged less than 50 years: an Australian case-control study. Cancer causes & control: CCC. 2010;21:2303–2308. - PubMed
-
- Tuckerman EM, Okon MA, Li T, Laird SM. Do androgens have a direct effect on endometrial function? An in vitro study. Fertil Steril. 2000;74:771–779. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
