

Vibrio fischeri-derived outer membrane vesicles trigger host development
- PMID: 26399913
- PMCID: PMC4803540
- DOI: 10.1111/cmi.12525
Vibrio fischeri-derived outer membrane vesicles trigger host development
Abstract
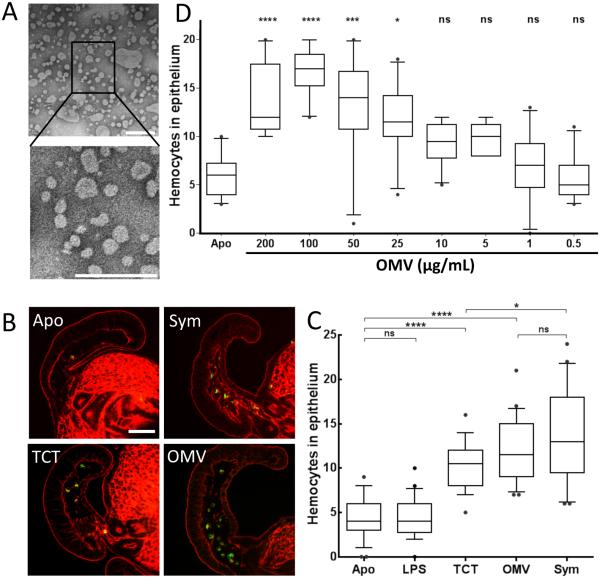
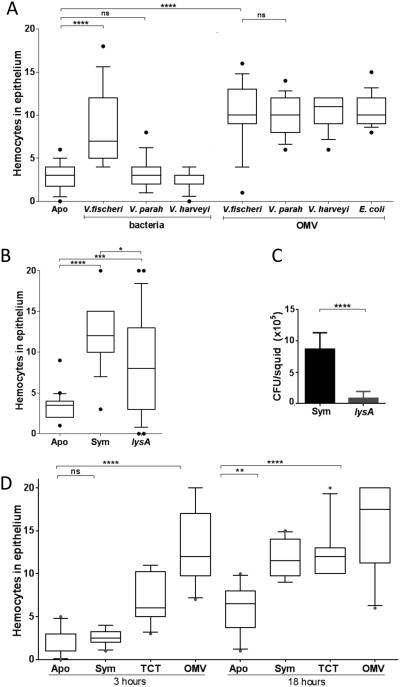
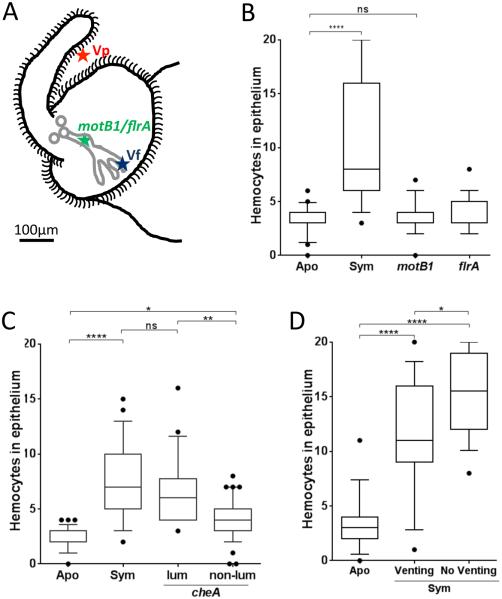
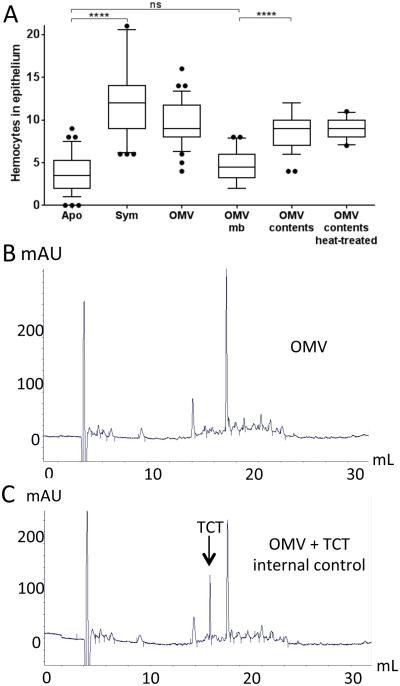
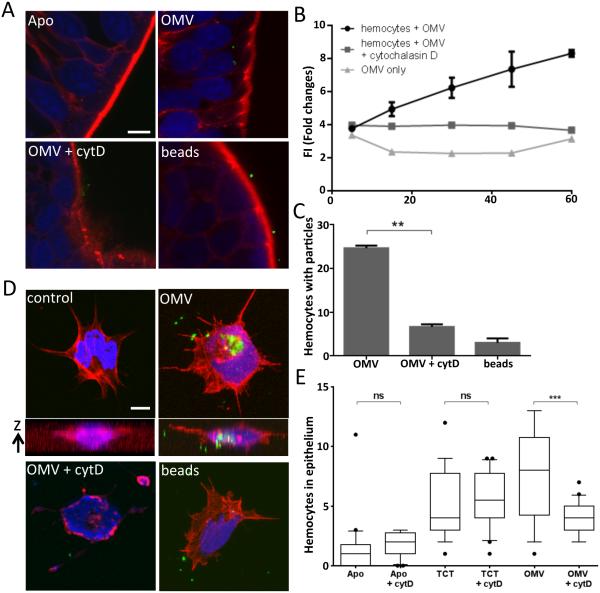
Outer membrane vesicles (OMV) are critical elements in many host-cell/microbe interactions. Previous studies of the symbiotic association between Euprymna scolopes and Vibrio fischeri had shown that within 12 h of colonizing crypts deep within the squid's light organ, the symbionts trigger an irreversible programme of tissue development in the host. Here, we report that OMV produced by V. fischeri are powerful contributors to this process. The first detectable host response to the OMV is an increased trafficking of macrophage-like cells called haemocytes into surface epithelial tissues. We showed that exposing the squid to other Vibrio species fails to induce this trafficking; however, addition of a high concentration of their OMV, which can diffuse into the crypts, does. We also provide evidence that tracheal cytotoxin released by the symbionts, which can induce haemocyte trafficking, is not part of the OMV cargo, suggesting two distinct mechanisms to induce the same morphogenesis event. By manipulating the timing and localization of OMV signal delivery, we showed that haemocyte trafficking is fully induced only when V. fischeri, the sole species able to reach and grow in the crypts, succeeds in establishing a sustained colonization. Further, our data suggest that the host's detection of OMV serves as a symbiotic checkpoint prior to inducing irreversible morphogenesis.
Keywords: OMV; euprymna scolopes; haemocyte; immune response; symbiosis.
© 2015 John Wiley & Sons Ltd.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
