

Definition of Proteasomal Peptide Splicing Rules for High-Efficiency Spliced Peptide Presentation by MHC Class I Molecules
- PMID: 26401003
- PMCID: PMC4642839
- DOI: 10.4049/jimmunol.1402455
Definition of Proteasomal Peptide Splicing Rules for High-Efficiency Spliced Peptide Presentation by MHC Class I Molecules
Abstract
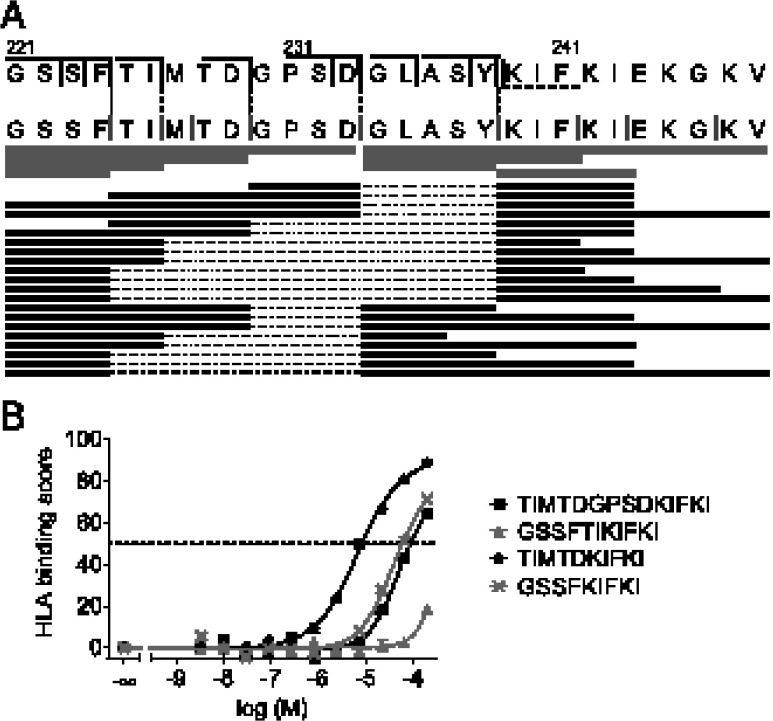
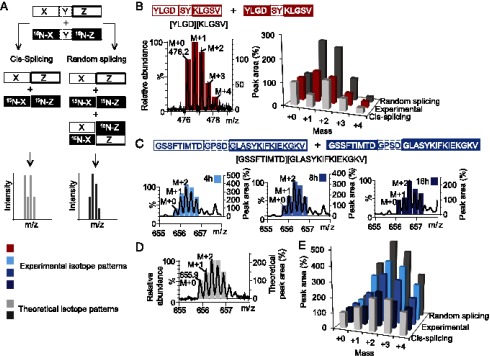
Peptide splicing, in which two distant parts of a protein are excised and then ligated to form a novel peptide, can generate unique MHC class I-restricted responses. Because these peptides are not genetically encoded and the rules behind proteasomal splicing are unknown, it is difficult to predict these spliced Ags. In the current study, small libraries of short peptides were used to identify amino acid sequences that affect the efficiency of this transpeptidation process. We observed that splicing does not occur at random, neither in terms of the amino acid sequences nor through random splicing of peptides from different sources. In contrast, splicing followed distinct rules that we deduced and validated both in vitro and in cells. Peptide ligation was quantified using a model peptide and demonstrated to occur with up to 30% ligation efficiency in vitro, provided that optimal structural requirements for ligation were met by both ligating partners. In addition, many splicing products could be formed from a single protein. Our splicing rules will facilitate prediction and detection of new spliced Ags to expand the peptidome presented by MHC class I Ags.
Copyright © 2015 by The American Association of Immunologists, Inc.
Figures


References
-
- Neefjes J., Jongsma M. L., Paul P., Bakke O. 2011. Towards a systems understanding of MHC class I and MHC class II antigen presentation. Nat. Rev. Immunol. 11: 823–836. - PubMed
-
- Vigneron N., Van den Eynde B. J. 2012. Proteasome subtypes and the processing of tumor antigens: increasing antigenic diversity. Curr. Opin. Immunol. 24: 84–91. - PubMed
-
- Basler M., Kirk C. J., Groettrup M. 2013. The immunoproteasome in antigen processing and other immunological functions. Curr. Opin. Immunol. 25: 74–80. - PubMed
-
- Vigneron N., Stroobant V., Chapiro J., Ooms A., Degiovanni G., Morel S., van der Bruggen P., Boon T., Van den Eynde B. J. 2004. An antigenic peptide produced by peptide splicing in the proteasome. Science 304: 587–590. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
