

Different Stress-Induced Calcium Signatures Are Reported by Aequorin-Mediated Calcium Measurements in Living Cells of Aspergillus fumigatus
- PMID: 26402916
- PMCID: PMC4581630
- DOI: 10.1371/journal.pone.0138008
Different Stress-Induced Calcium Signatures Are Reported by Aequorin-Mediated Calcium Measurements in Living Cells of Aspergillus fumigatus
Abstract
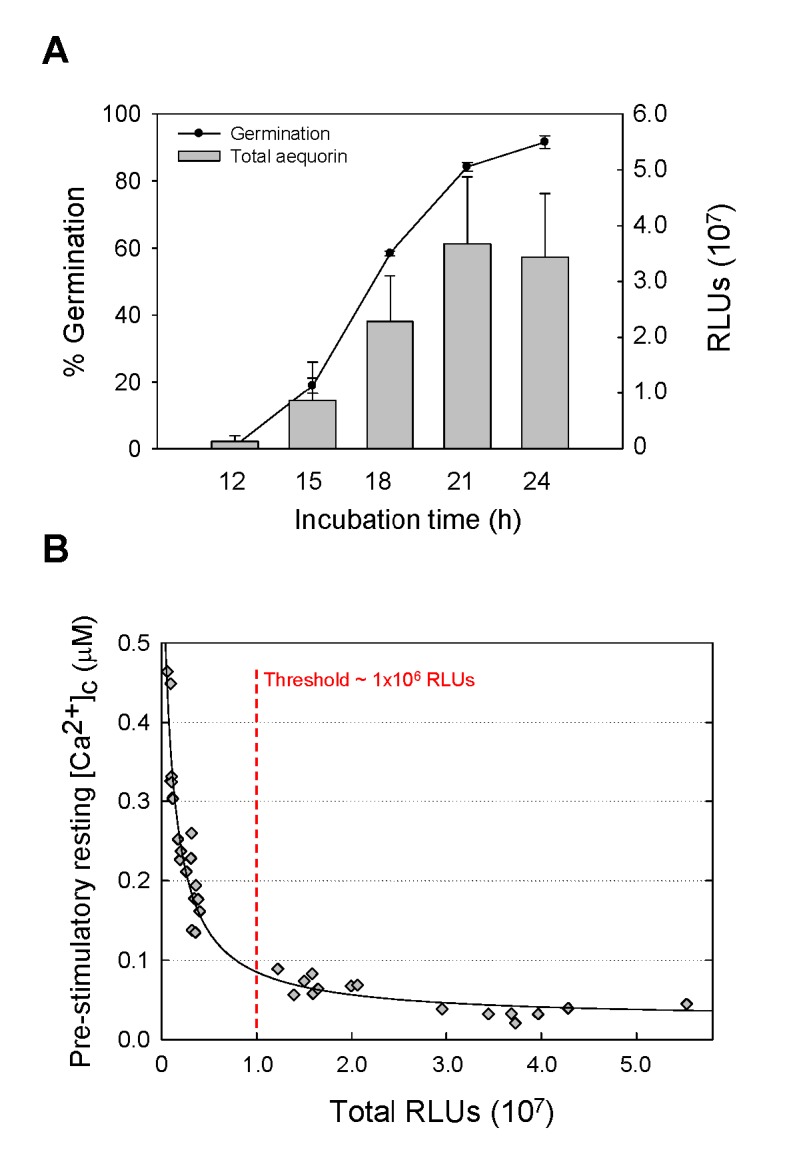
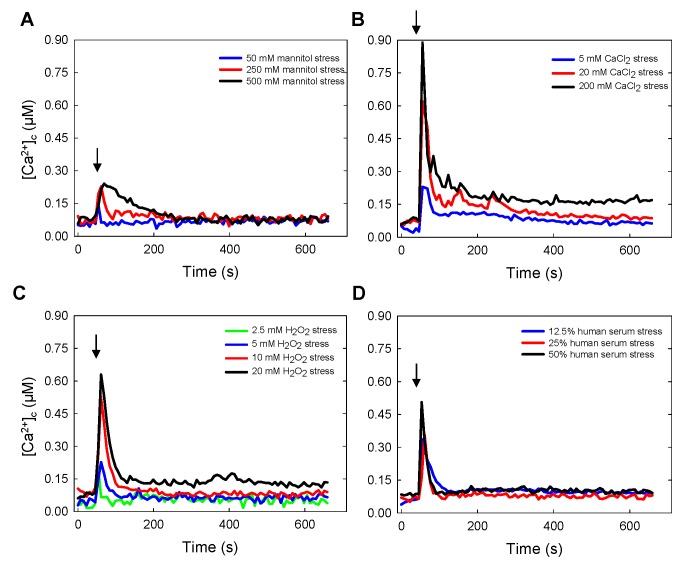
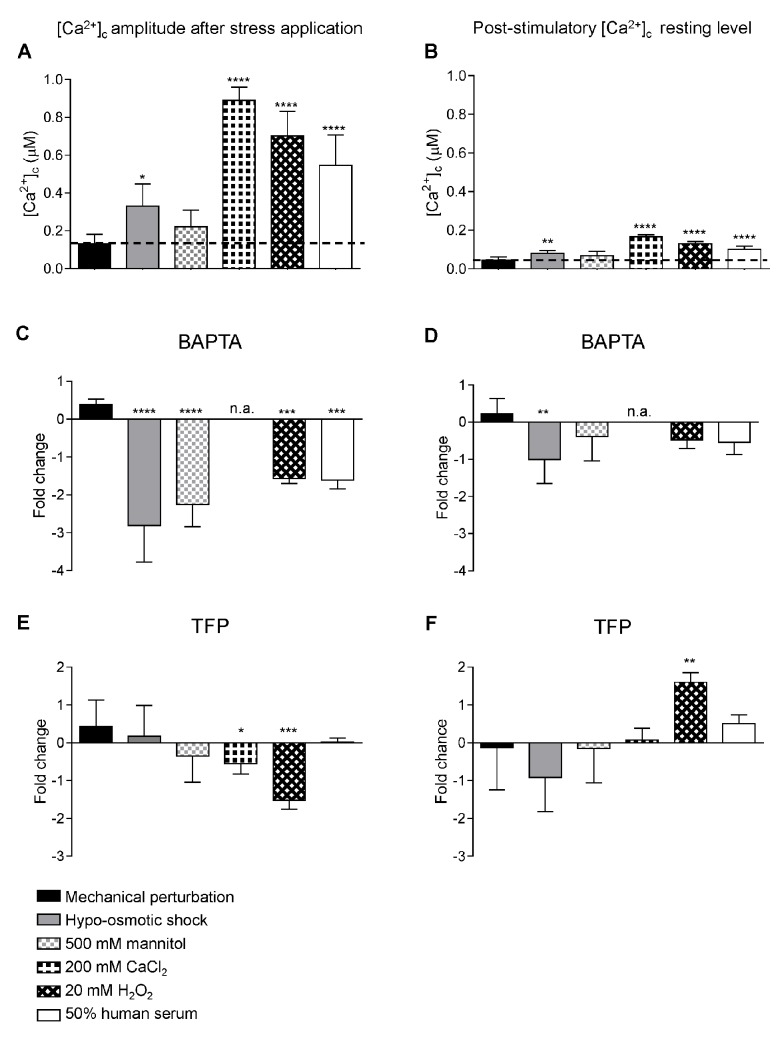
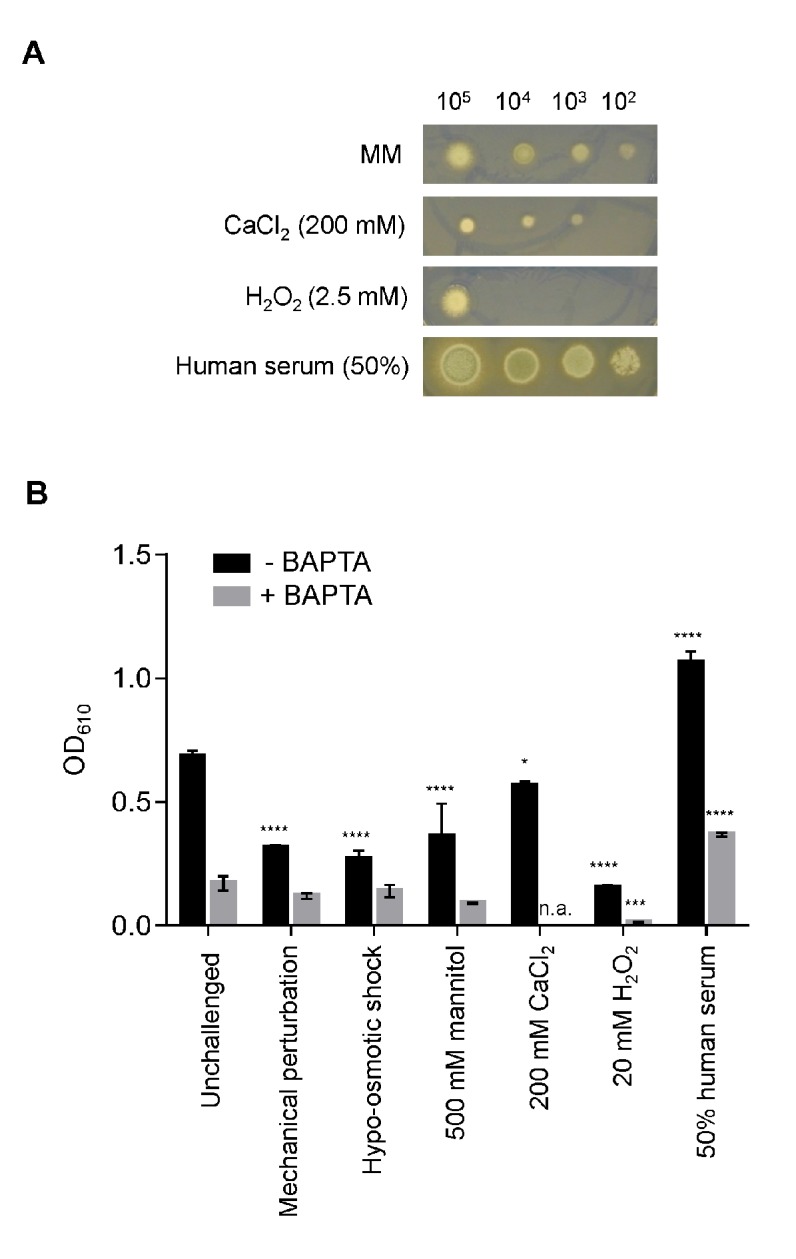
Aspergillus fumigatus is an inhaled fungal pathogen of human lungs, the developmental growth of which is reliant upon Ca2+-mediated signalling. Ca2+ signalling has regulatory significance in all eukaryotic cells but how A. fumigatus uses intracellular Ca2+ signals to respond to stresses imposed by the mammalian lung is poorly understood. In this work, A. fumigatus strains derived from the clinical isolate CEA10, and a non-homologous recombination mutant ΔakuBKU80, were engineered to express the bioluminescent Ca2+-reporter aequorin. An aequorin-mediated method for routine Ca2+ measurements during the early stages of colony initiation was successfully developed and dynamic changes in cytosolic free calcium ([Ca2+]c) in response to extracellular stimuli were measured. The response to extracellular challenges (hypo- and hyper-osmotic shock, mechanical perturbation, high extracellular Ca2+, oxidative stress or exposure to human serum) that the fungus might be exposed to during infection, were analysed in living conidial germlings. The 'signatures' of the transient [Ca2+]c responses to extracellular stimuli were found to be dose- and age-dependent. Moreover, Ca2+-signatures associated with each physico-chemical treatment were found to be unique, suggesting the involvement of heterogeneous combinations of Ca2+-signalling components in each stress response. Concordant with the involvement of Ca2+-calmodulin complexes in these Ca2+-mediated responses, the calmodulin inhibitor trifluoperazine (TFP) induced changes in the Ca2+-signatures to all the challenges. The Ca2+-chelator BAPTA potently inhibited the initial responses to most stressors in accordance with a critical role for extracellular Ca2+ in initiating the stress responses.
Conflict of interest statement
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
