

PTEN regulates RPA1 and protects DNA replication forks
- PMID: 26403191
- PMCID: PMC4650420
- DOI: 10.1038/cr.2015.115
PTEN regulates RPA1 and protects DNA replication forks
Abstract
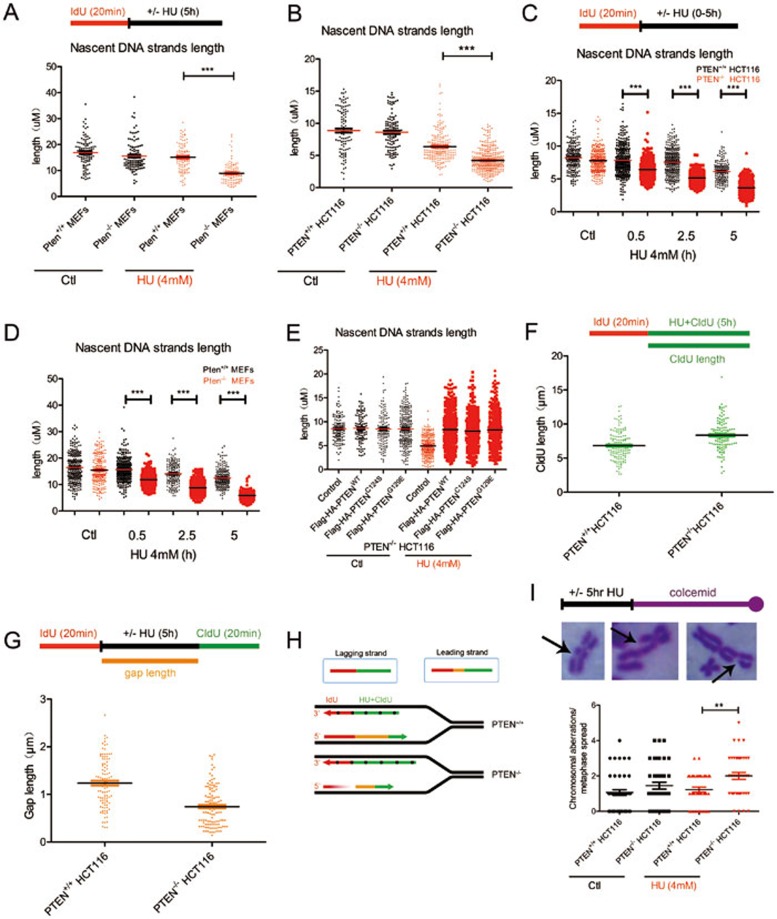
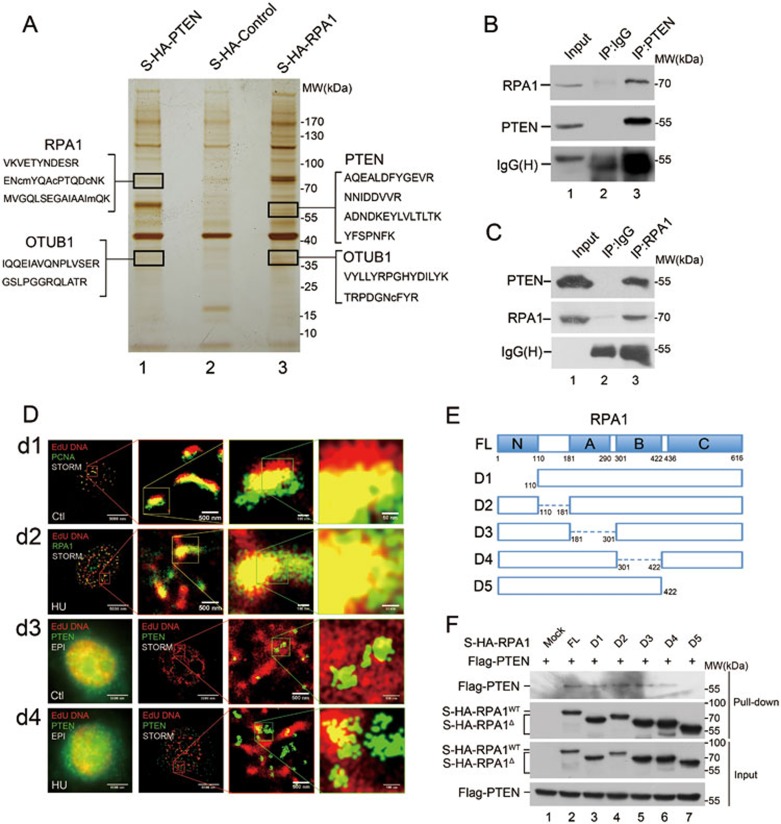
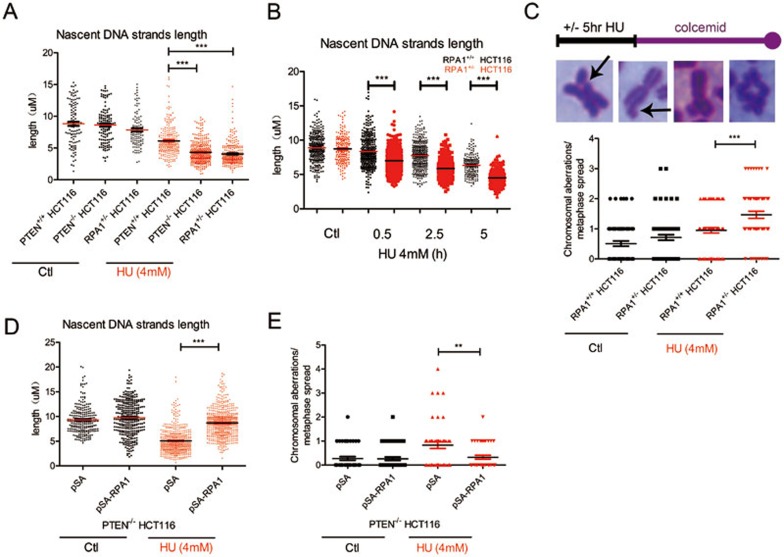
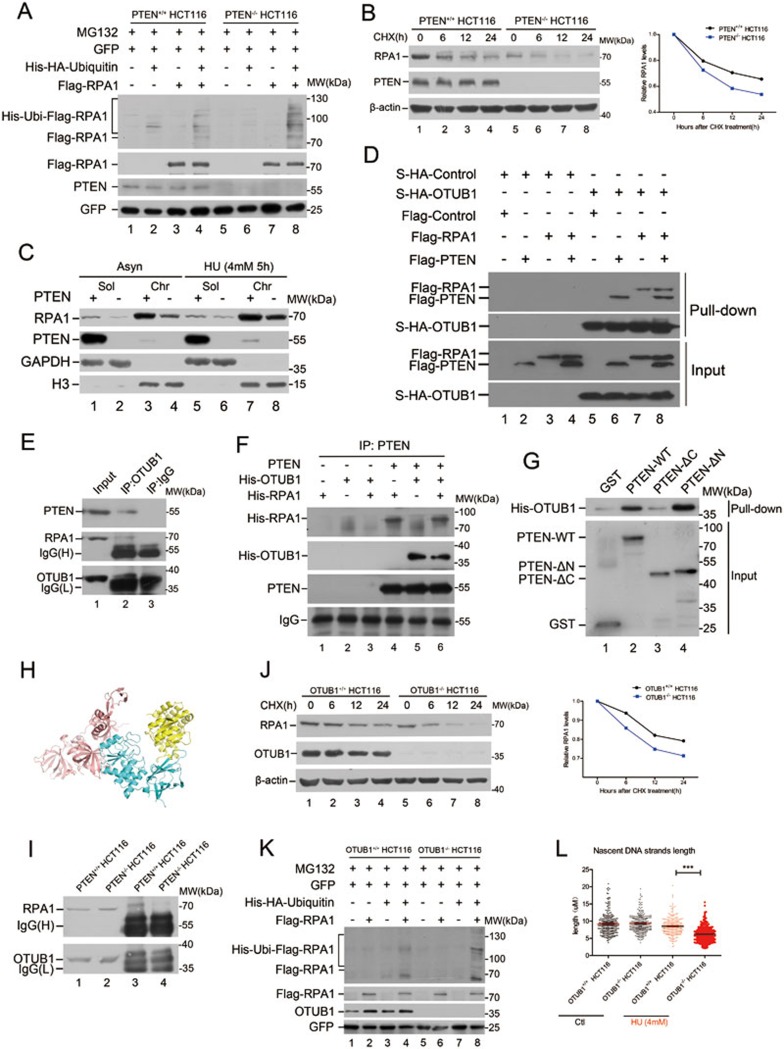
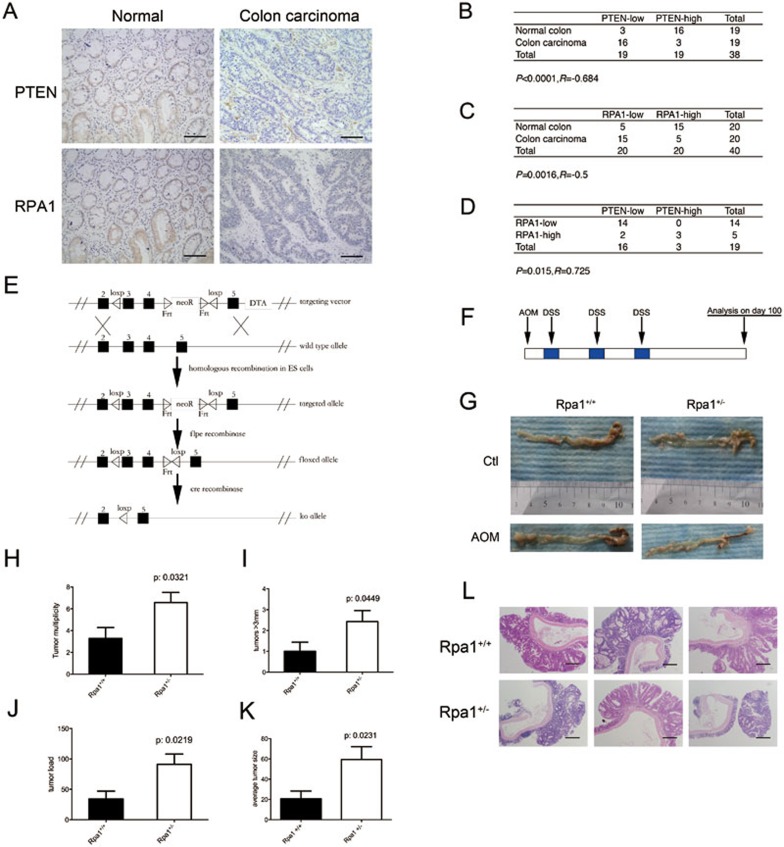
Tumor suppressor PTEN regulates cellular activities and controls genome stability through multiple mechanisms. In this study, we report that PTEN is necessary for the protection of DNA replication forks against replication stress. We show that deletion of PTEN leads to replication fork collapse and chromosomal instability upon fork stalling following nucleotide depletion induced by hydroxyurea. PTEN is physically associated with replication protein A 1 (RPA1) via the RPA1 C-terminal domain. STORM and iPOND reveal that PTEN is localized at replication sites and promotes RPA1 accumulation on replication forks. PTEN recruits the deubiquitinase OTUB1 to mediate RPA1 deubiquitination. RPA1 deletion confers a phenotype like that observed in PTEN knockout cells with stalling of replication forks. Expression of PTEN and RPA1 shows strong correlation in colorectal cancer. Heterozygous disruption of RPA1 promotes tumorigenesis in mice. These results demonstrate that PTEN is essential for DNA replication fork protection. We propose that RPA1 is a target of PTEN function in fork protection and that PTEN maintains genome stability through regulation of DNA replication.
Figures
Comment on
-
Soluble protein oligomers in neurodegeneration: lessons from the Alzheimer's amyloid beta-peptide.Nat Rev Mol Cell Biol. 2007 Feb;8(2):101-12. doi: 10.1038/nrm2101. Nat Rev Mol Cell Biol. 2007. PMID: 17245412 Review.
Similar articles
-
PTEN in the maintenance of genome integrity: From DNA replication to chromosome segregation.Bioessays. 2017 Oct;39(10):10.1002/bies.201700082. doi: 10.1002/bies.201700082. Epub 2017 Sep 11. Bioessays. 2017. PMID: 28891157 Free PMC article. Review.
-
In and out of Replication Stress: PCNA/RPA1-Based Dynamics of Fork Stalling and Restart in the Same Cell.Int J Mol Sci. 2025 Jan 14;26(2):667. doi: 10.3390/ijms26020667. Int J Mol Sci. 2025. PMID: 39859385 Free PMC article.
-
PTEN Controls the DNA Replication Process through MCM2 in Response to Replicative Stress.Cell Rep. 2015 Nov 17;13(7):1295-1303. doi: 10.1016/j.celrep.2015.10.016. Epub 2015 Nov 5. Cell Rep. 2015. PMID: 26549452
-
Deletion of BRCA2 exon 27 causes defects in response to both stalled and collapsed replication forks.Mutat Res. 2014 Aug-Sep;766-767:66-72. doi: 10.1016/j.mrfmmm.2014.06.003. Epub 2014 Jun 22. Mutat Res. 2014. PMID: 25847274
-
Preventing replication fork collapse to maintain genome integrity.DNA Repair (Amst). 2015 Aug;32:149-157. doi: 10.1016/j.dnarep.2015.04.026. Epub 2015 May 1. DNA Repair (Amst). 2015. PMID: 25957489 Free PMC article. Review.
Cited by
-
Targeting phosphomevalonate kinase enhances radiosensitivity via ubiquitination of the replication protein A1 in lung cancer cells.Cancer Sci. 2023 Sep;114(9):3583-3594. doi: 10.1111/cas.15896. Epub 2023 Jul 5. Cancer Sci. 2023. PMID: 37650703 Free PMC article.
-
PTEN in the maintenance of genome integrity: From DNA replication to chromosome segregation.Bioessays. 2017 Oct;39(10):10.1002/bies.201700082. doi: 10.1002/bies.201700082. Epub 2017 Sep 11. Bioessays. 2017. PMID: 28891157 Free PMC article. Review.
-
Global crotonylome reveals CDYL-regulated RPA1 crotonylation in homologous recombination-mediated DNA repair.Sci Adv. 2020 Mar 13;6(11):eaay4697. doi: 10.1126/sciadv.aay4697. eCollection 2020 Mar. Sci Adv. 2020. PMID: 32201722 Free PMC article.
-
The Mechanisms of lncRNA-Mediated Multidrug Resistance and the Clinical Application Prospects of lncRNAs in Breast Cancer.Cancers (Basel). 2022 Apr 23;14(9):2101. doi: 10.3390/cancers14092101. Cancers (Basel). 2022. PMID: 35565231 Free PMC article. Review.
-
PARP inhibitor resistance: the underlying mechanisms and clinical implications.Mol Cancer. 2020 Jun 20;19(1):107. doi: 10.1186/s12943-020-01227-0. Mol Cancer. 2020. PMID: 32563252 Free PMC article. Review.
References
-
- 1Aguilera A, Gomez-Gonzalez B. Genome instability: a mechanistic view of its causes and consequences. Nat Rev Genet 2008; 9:204–217. - PubMed
-
- 2Branzei D, Foiani M. Maintaining genome stability at the replication fork. Nat Rev Mol Cell Biol 2010; 11:208–219. - PubMed
-
- 4Negrini S, Gorgoulis VG, Halazonetis TD. Genomic instability--an evolving hallmark of cancer. Nat Rev Mol Cell Biol 2010; 11:220–228. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
