

The importance of the Non Obese Diabetic (NOD) mouse model in autoimmune diabetes
- PMID: 26403950
- PMCID: PMC4765310
- DOI: 10.1016/j.jaut.2015.08.019
The importance of the Non Obese Diabetic (NOD) mouse model in autoimmune diabetes
Abstract
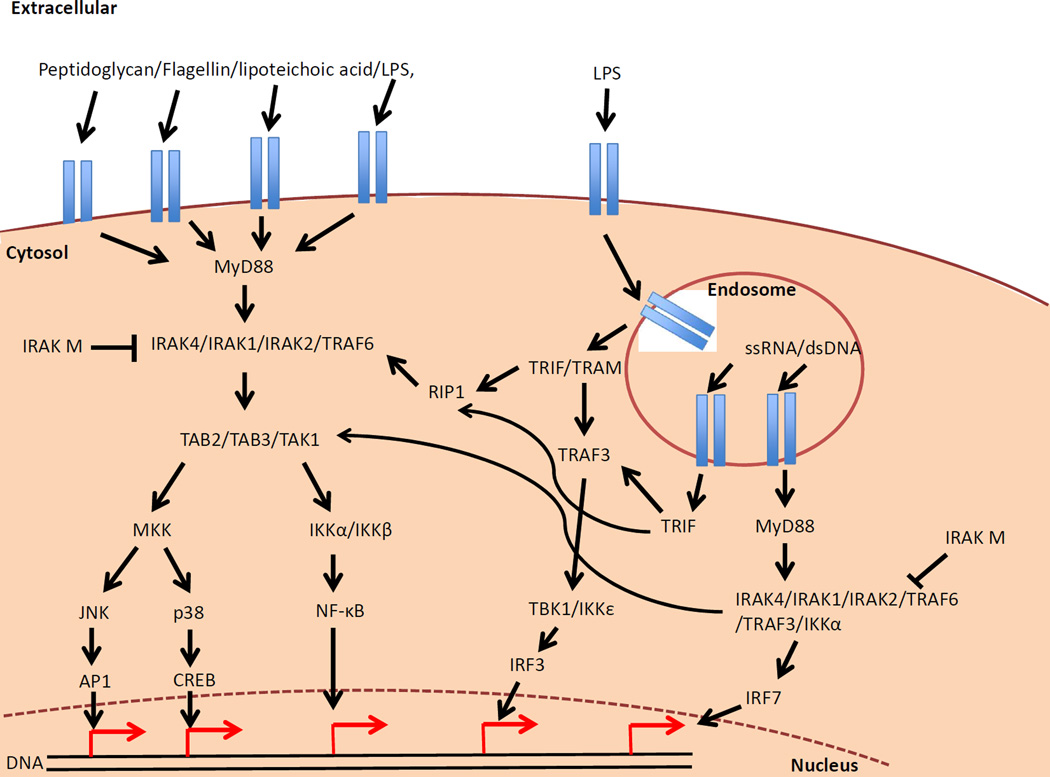
Type 1 Diabetes (T1D) is an autoimmune disease characterized by the pancreatic infiltration of immune cells resulting in T cell-mediated destruction of the insulin-producing beta cells. The successes of the Non-Obese Diabetic (NOD) mouse model have come in multiple forms including identifying key genetic and environmental risk factors e.g. Idd loci and effects of microorganisms including the gut microbiota, respectively, and how they may contribute to disease susceptibility and pathogenesis. Furthermore, the NOD model also provides insights into the roles of the innate immune cells as well as the B cells in contributing to the T cell-mediated disease. Unlike many autoimmune disease models, the NOD mouse develops spontaneous disease and has many similarities to human T1D. Through exploiting these similarities many targets have been identified for immune-intervention strategies. Although many of these immunotherapies did not have a significant impact on human T1D, they have been shown to be effective in the NOD mouse in early stage disease, which is not equivalent to trials in newly-diagnosed patients with diabetes. However, the continued development of humanized NOD mice would enable further clinical developments, bringing T1D research to a new translational level. Therefore, it is the aim of this review to discuss the importance of the NOD model in identifying the roles of the innate immune system and the interaction with the gut microbiota in modifying diabetes susceptibility. In addition, the role of the B cells will also be discussed with new insights gained through B cell depletion experiments and the impact on translational developments. Finally, this review will also discuss the future of the NOD mouse and the development of humanized NOD mice, providing novel insights into human T1D.
Keywords: B cells; Gut microbiota; Humanized mice; NOD; Type 1 diabetes.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures


References
-
- Gillespie KM, Bain SC, Barnett AH, Bingley PJ, Christie MR, Gill GV, et al. The rising incidence of childhood type 1 diabetes and reduced contribution of high-risk HLA haplotypes. Lancet. 2004;364:1699–1700. - PubMed
-
- Bessaoud K, Boudraa G, Molinero de Ropolo M, de Sereday M, Marti ML, Moser M, et al. Incidence and trends of childhood Type 1 diabetes worldwide 1990–1999. Diabetic Medicine. 2006;23:857–866. - PubMed
-
- Harjutsalo V, Sjöberg L, Tuomilehto J. Time trends in the incidence of type 1 diabetes in Finnish children: a cohort study. Lancet. 2008;371:1777–1782. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P30-DK-45735/DK/NIDDK NIH HHS/United States
- MR/K021141/MRC_/Medical Research Council/United Kingdom
- RC1 DK087699/DK/NIDDK NIH HHS/United States
- 08/0003767/DUK_/Diabetes UK/United Kingdom
- R01 DK088181/DK/NIDDK NIH HHS/United States
- G0901155/MRC_/Medical Research Council/United Kingdom
- P30 DK045735/DK/NIDDK NIH HHS/United States
- DK088181/DK/NIDDK NIH HHS/United States
- R01 DK092882/DK/NIDDK NIH HHS/United States
- R01 DK100500/DK/NIDDK NIH HHS/United States
- DK092882/DK/NIDDK NIH HHS/United States
- RC1DK-087699/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
