

Mechanisms of stress in the brain
- PMID: 26404710
- PMCID: PMC4933289
- DOI: 10.1038/nn.4086
Mechanisms of stress in the brain
Abstract
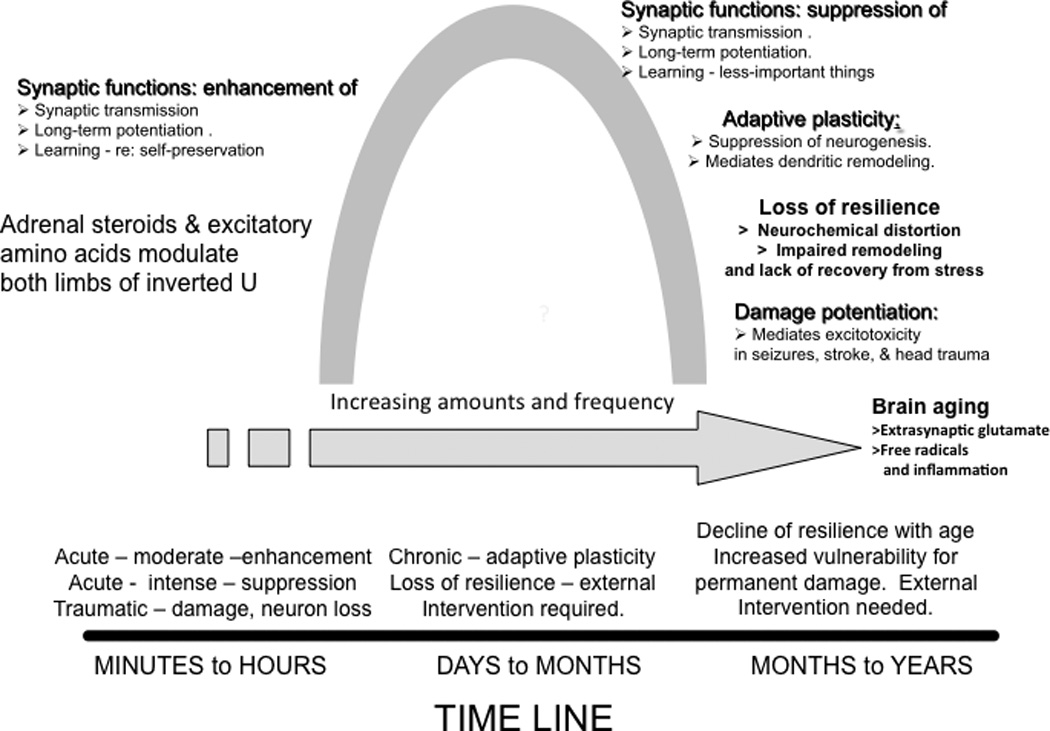
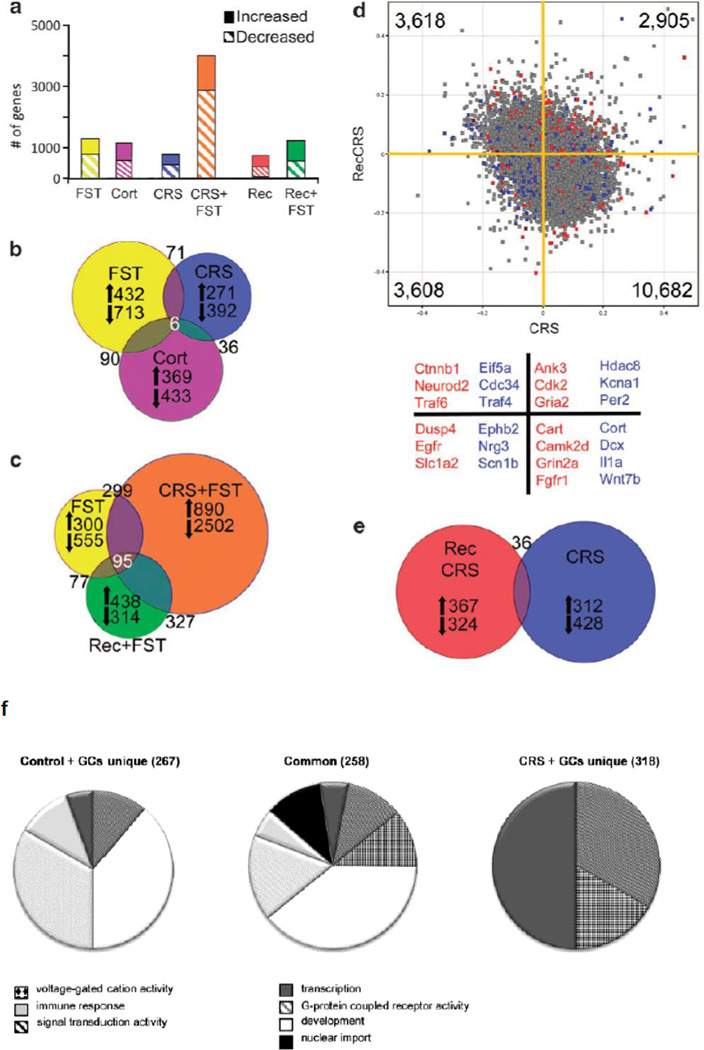
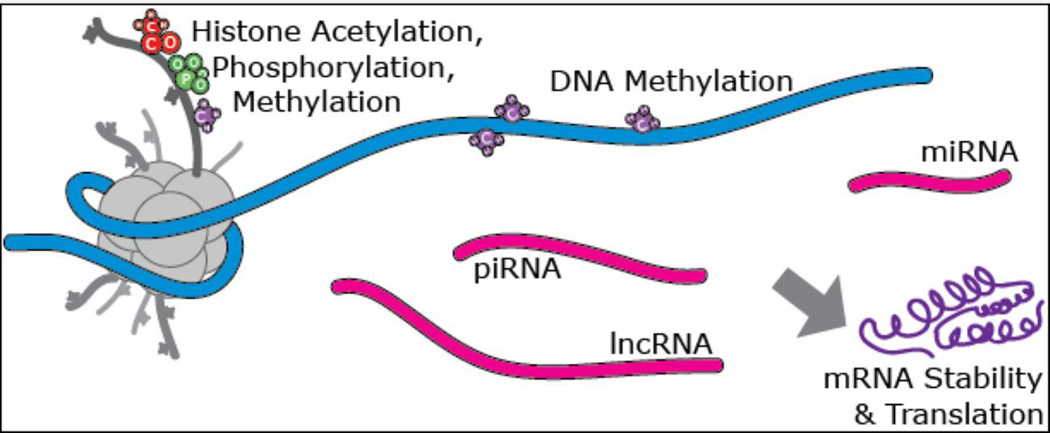
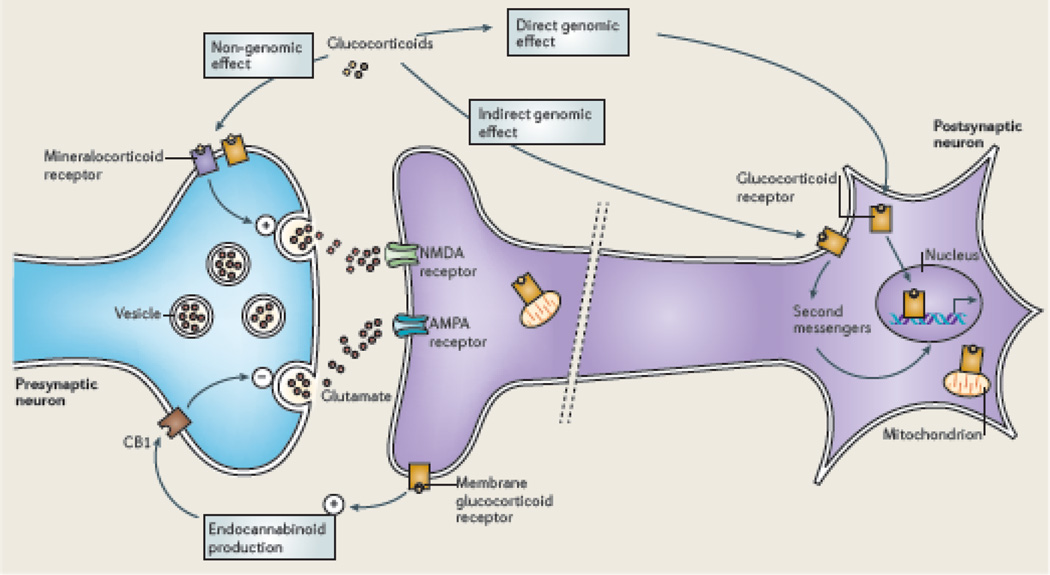
The brain is the central organ involved in perceiving and adapting to social and physical stressors via multiple interacting mediators, from the cell surface to the cytoskeleton to epigenetic regulation and nongenomic mechanisms. A key result of stress is structural remodeling of neural architecture, which may be a sign of successful adaptation, whereas persistence of these changes when stress ends indicates failed resilience. Excitatory amino acids and glucocorticoids have key roles in these processes, along with a growing list of extra- and intracellular mediators that includes endocannabinoids and brain-derived neurotrophic factor (BDNF). The result is a continually changing pattern of gene expression mediated by epigenetic mechanisms involving histone modifications and CpG methylation and hydroxymethylation as well as by the activity of retrotransposons that may alter genomic stability. Elucidation of the underlying mechanisms of plasticity and vulnerability of the brain provides a basis for understanding the efficacy of interventions for anxiety and depressive disorders as well as age-related cognitive decline.
Conflict of interest statement
The authors declare that there are no conflicts of interest.
Figures
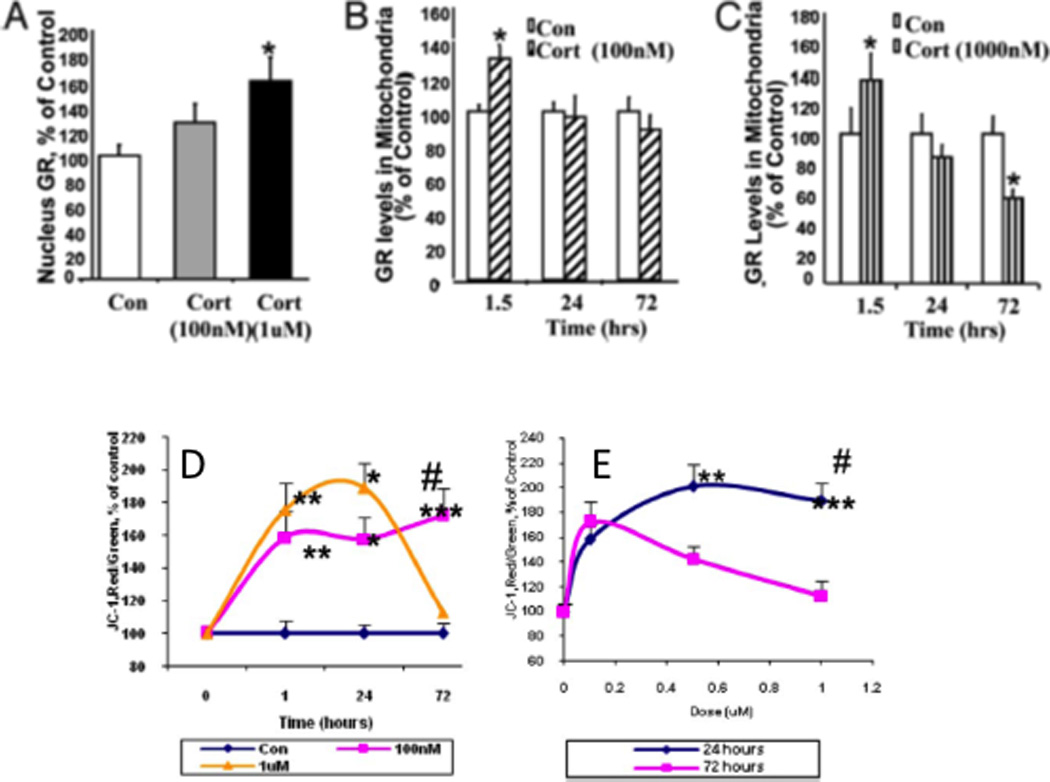
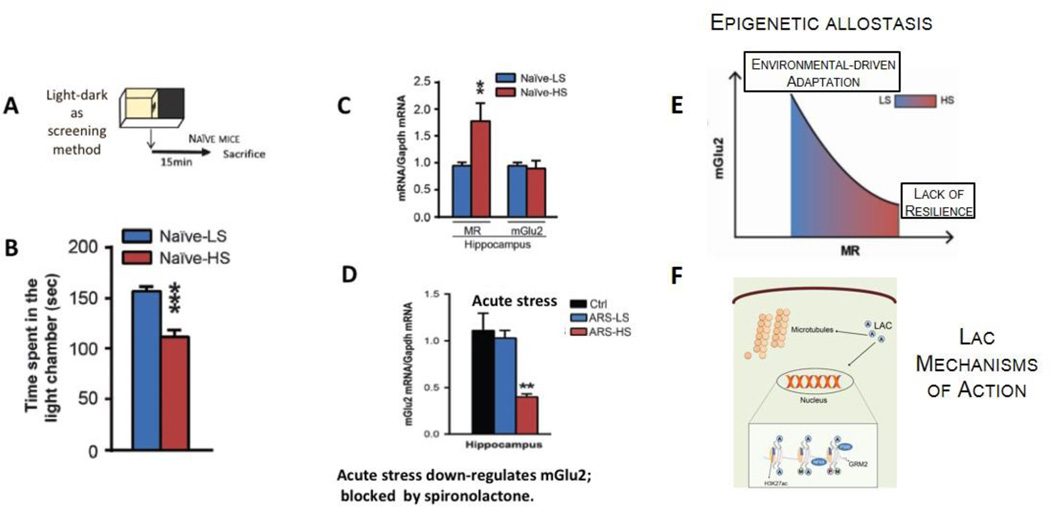
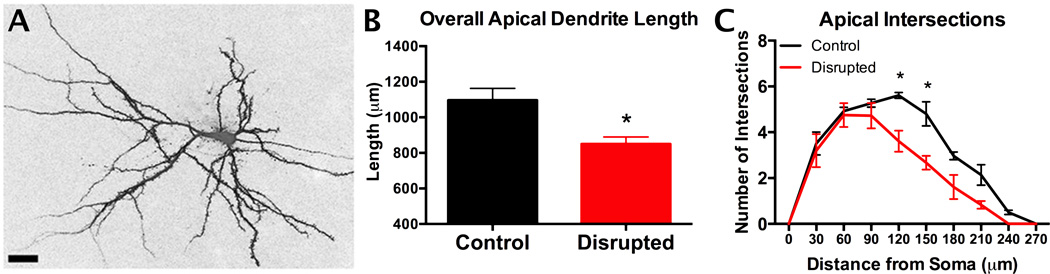
References
-
- McEwen BS. Protective and Damaging Effects of Stress Mediators. New England J. Med. 1998;338:171–179. - PubMed
-
- McEwen BS, Wingfield JC. The concept of allostasis in biology and biomedicine. Horm. & Behav. 2003;43:2–15. - PubMed
-
- Shonkoff JP, Boyce WT, McEwen BS. Neuroscience, molecular biology, and the childhood roots of health disparities. JAMA. 2009;301:2252–2259. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
