

RIPK1 and NF-κB signaling in dying cells determines cross-priming of CD8⁺ T cells
- PMID: 26405229
- PMCID: PMC4651449
- DOI: 10.1126/science.aad0395
RIPK1 and NF-κB signaling in dying cells determines cross-priming of CD8⁺ T cells
Abstract
Dying cells initiate adaptive immunity by providing both antigens and inflammatory stimuli for dendritic cells, which in turn activate CD8(+) T cells through a process called antigen cross-priming. To define how different forms of programmed cell death influence immunity, we established models of necroptosis and apoptosis, in which dying cells are generated by receptor-interacting protein kinase-3 and caspase-8 dimerization, respectively. We found that the release of inflammatory mediators, such as damage-associated molecular patterns, by dying cells was not sufficient for CD8(+) T cell cross-priming. Instead, robust cross-priming required receptor-interacting protein kinase-1 (RIPK1) signaling and nuclear factor κB (NF-κB)-induced transcription within dying cells. Decoupling NF-κB signaling from necroptosis or inflammatory apoptosis reduced priming efficiency and tumor immunity. Our results reveal that coordinated inflammatory and cell death signaling pathways within dying cells orchestrate adaptive immunity.
Copyright © 2015, American Association for the Advancement of Science.
Figures
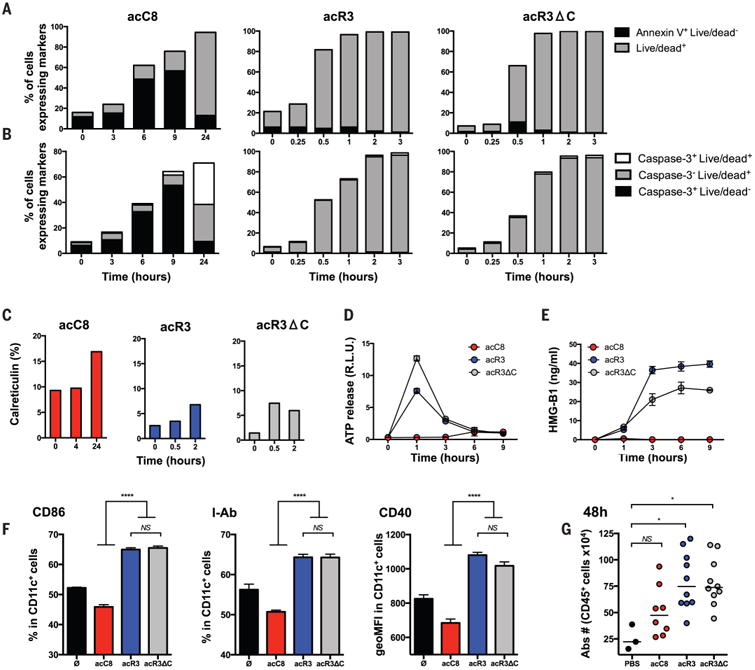
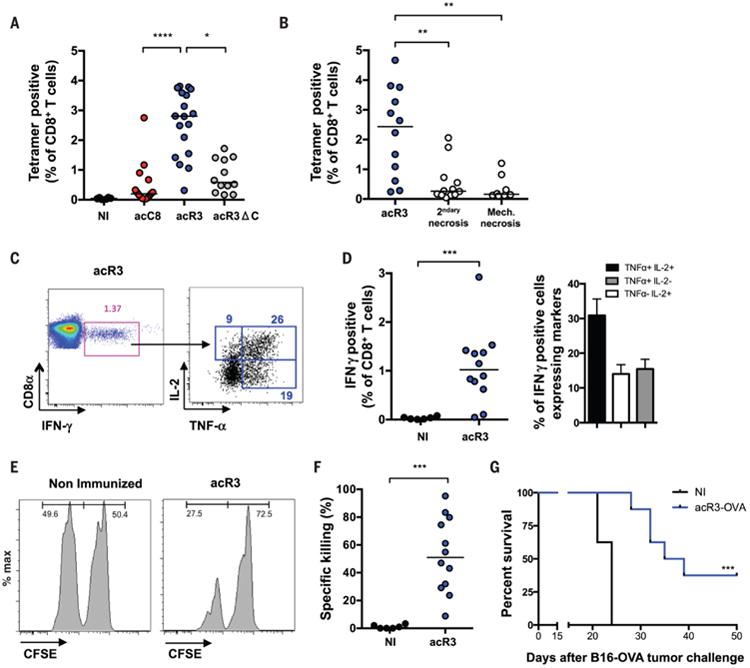
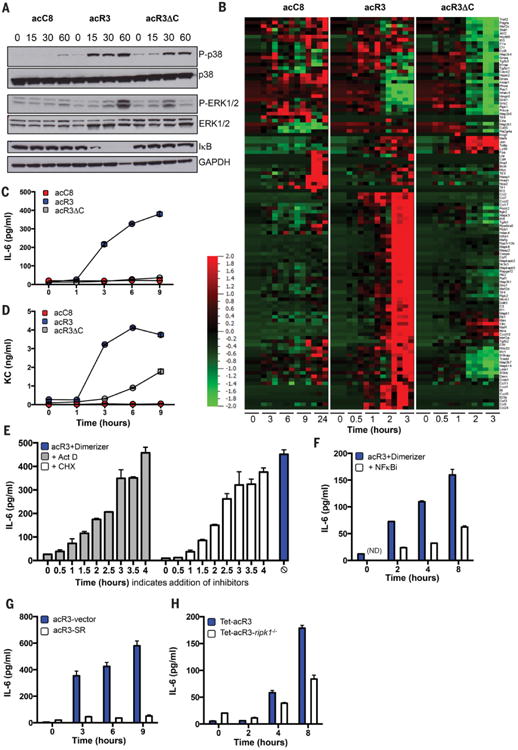
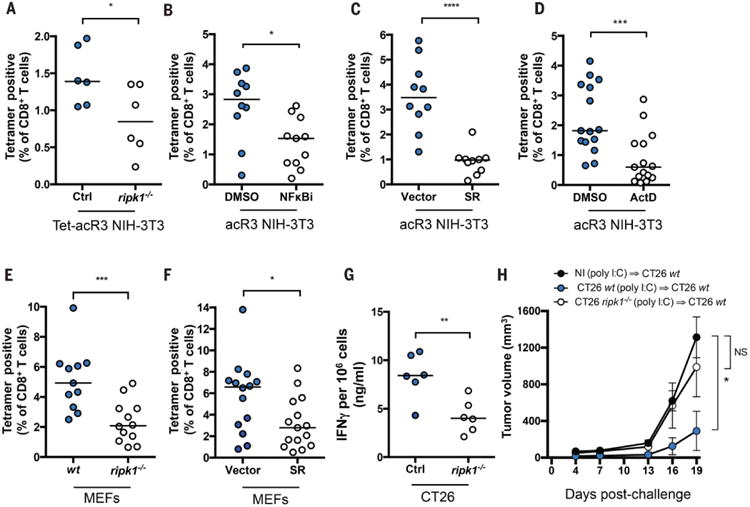
Comment in
-
Cell death: Pathways for cross-priming.Nat Rev Immunol. 2015 Dec;15(12):725. doi: 10.1038/nri3933. Epub 2015 Oct 23. Nat Rev Immunol. 2015. PMID: 26493941 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
