

Plasmodium Infection Is Associated with Impaired Hepatic Dimethylarginine Dimethylaminohydrolase Activity and Disruption of Nitric Oxide Synthase Inhibitor/Substrate Homeostasis
- PMID: 26407009
- PMCID: PMC4583463
- DOI: 10.1371/journal.ppat.1005119
Plasmodium Infection Is Associated with Impaired Hepatic Dimethylarginine Dimethylaminohydrolase Activity and Disruption of Nitric Oxide Synthase Inhibitor/Substrate Homeostasis
Abstract
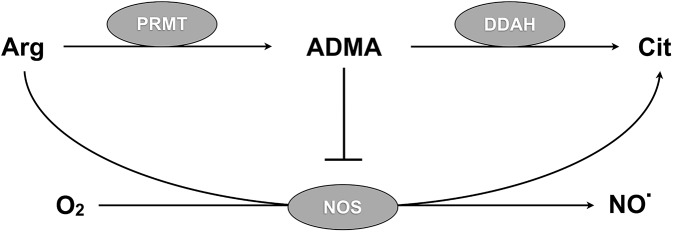
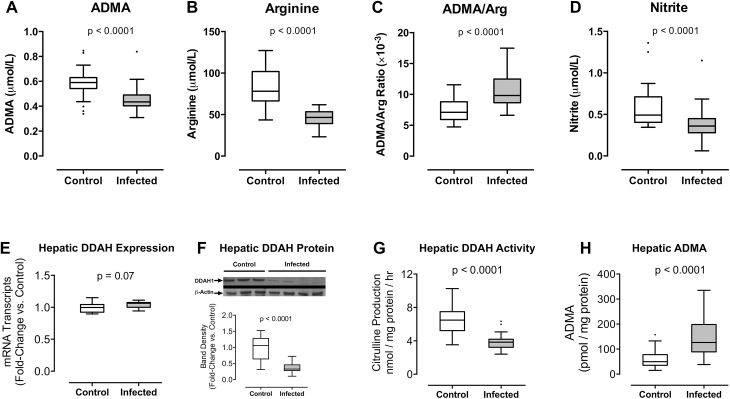
Inhibition of nitric oxide (NO) signaling may contribute to pathological activation of the vascular endothelium during severe malaria infection. Dimethylarginine dimethylaminohydrolase (DDAH) regulates endothelial NO synthesis by maintaining homeostasis between asymmetric dimethylarginine (ADMA), an endogenous NO synthase (NOS) inhibitor, and arginine, the NOS substrate. We carried out a community-based case-control study of Gambian children to determine whether ADMA and arginine homeostasis is disrupted during severe or uncomplicated malaria infections. Circulating plasma levels of ADMA and arginine were determined at initial presentation and 28 days later. Plasma ADMA/arginine ratios were elevated in children with acute severe malaria compared to 28-day follow-up values and compared to children with uncomplicated malaria or healthy children (p<0.0001 for each comparison). To test the hypothesis that DDAH1 is inactivated during Plasmodium infection, we examined DDAH1 in a mouse model of severe malaria. Plasmodium berghei ANKA infection inactivated hepatic DDAH1 via a post-transcriptional mechanism as evidenced by stable mRNA transcript number, decreased DDAH1 protein concentration, decreased enzyme activity, elevated tissue ADMA, elevated ADMA/arginine ratio in plasma, and decreased whole blood nitrite concentration. Loss of hepatic DDAH1 activity and disruption of ADMA/arginine homeostasis may contribute to severe malaria pathogenesis by inhibiting NO synthesis.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- World Health Organization. World malaria report 2014 [Internet]. 2014. Available: http://apps.who.int/iris/bitstream/10665/144852/2/9789241564830_eng.pdf?...
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
