

Adaptive Posttranslational Control in Cellular Stress Response Pathways and Its Relationship to Toxicity Testing and Safety Assessment
- PMID: 26408567
- PMCID: PMC4707198
- DOI: 10.1093/toxsci/kfv130
Adaptive Posttranslational Control in Cellular Stress Response Pathways and Its Relationship to Toxicity Testing and Safety Assessment
Abstract
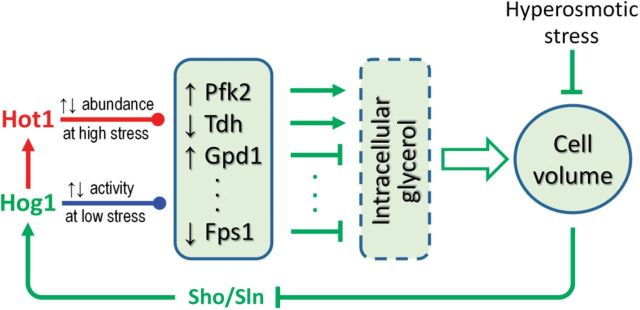
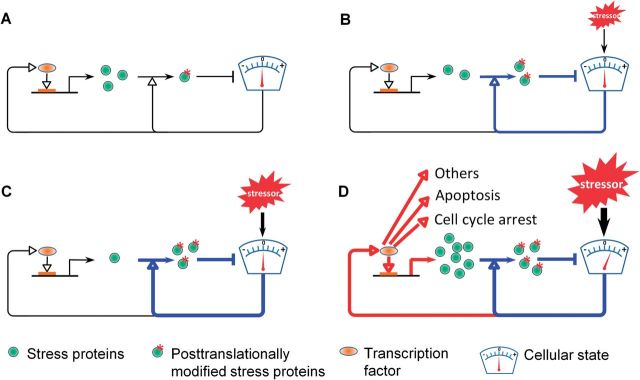
Although transcriptional induction of stress genes constitutes a major cellular defense program against a variety of stressors, posttranslational control directly regulating the activities of preexisting stress proteins provides a faster-acting alternative response. We propose that posttranslational control is a general adaptive mechanism operating in many stress pathways. Here with the aid of computational models, we first show that posttranslational control fulfills two roles: (1) handling small, transient stresses quickly and (2) stabilizing the negative feedback transcriptional network. We then review the posttranslational control pathways for major stress responses-oxidative stress, metal stress, hyperosmotic stress, DNA damage, heat shock, and hypoxia. Posttranslational regulation of stress protein activities occurs by reversible covalent modifications, allosteric or non-allosteric enzymatic regulations, and physically induced protein structural changes. Acting in feedback or feedforward networks, posttranslational control may establish a threshold level of cellular stress. Sub-threshold stresses are handled adequately by posttranslational control without invoking gene transcription. With supra-threshold stress levels, cellular homeostasis cannot be maintained and transcriptional induction of stress genes and other gene programs, eg, those regulating cell metabolism, proliferation, and apoptosis, takes place. The loss of homeostasis with consequent changes in cellular function may lead to adverse cellular outcomes. Overall, posttranslational and transcriptional control pathways constitute a stratified cellular defense system, handling stresses coherently across time and intensity. As cell-based assays become a focus for chemical testing anchored on toxicity pathways, examination of proteomic and metabolomic changes as a result of posttranslational control occurring in the absence of transcriptomic alterations deserves more attention.
Keywords: feedback; pathway; posttranslational; stress; transcriptional.
© The Author 2015. Published by Oxford University Press on behalf of the Society of Toxicology. All rights reserved. For Permissions, please e-mail: journals.permissions@oup.com.
Figures
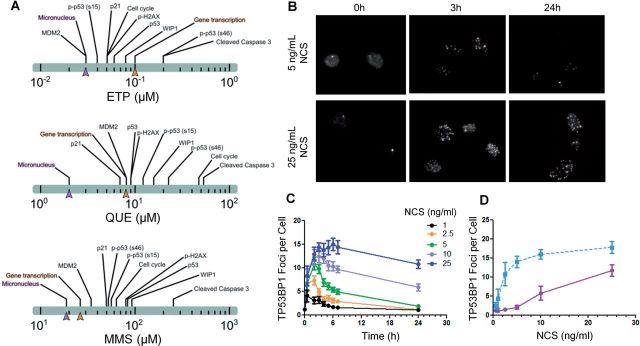
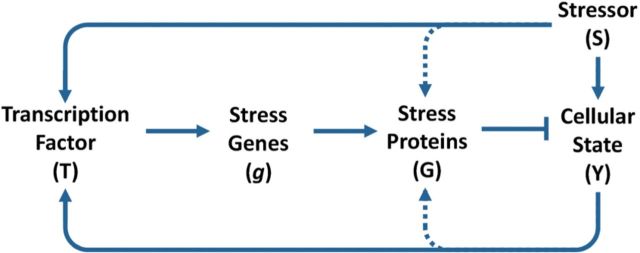
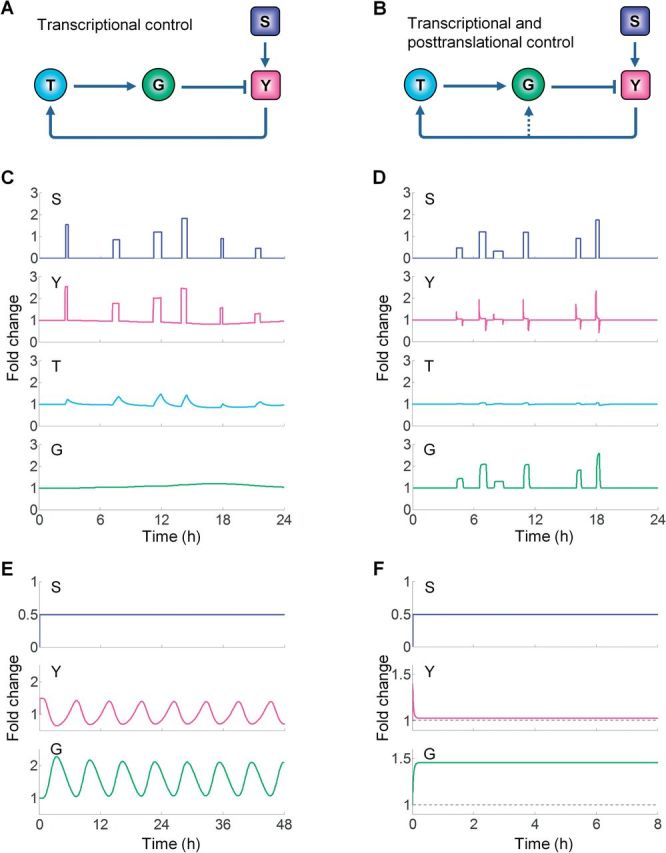
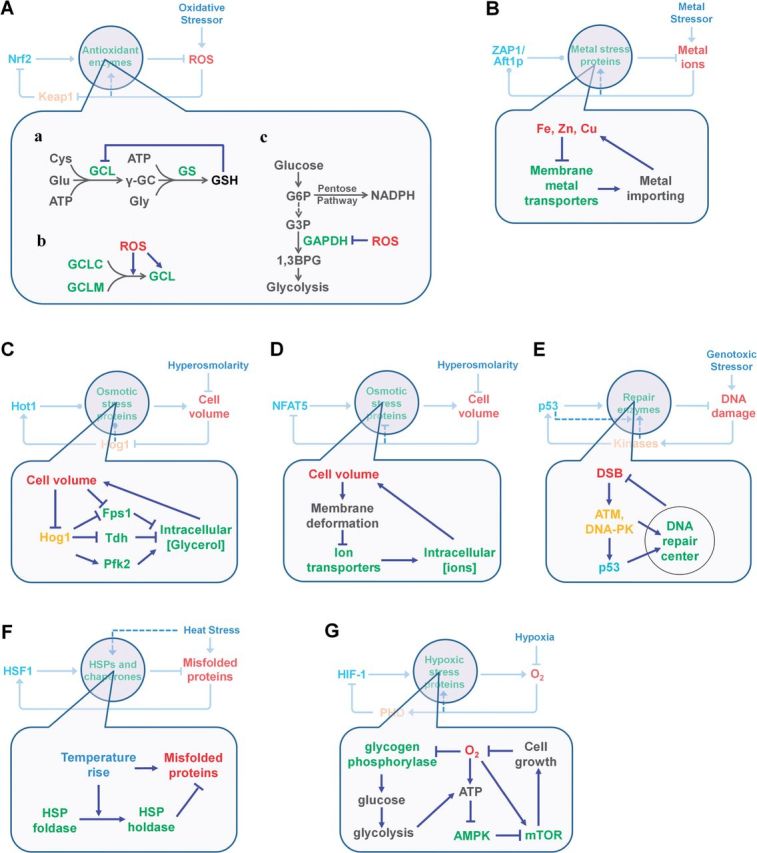
References
-
- Al Rashid S. T., Dellaire G., Cuddihy A., Jalali F., Vaid M., Coackley C., Folkard M., Xu Y., Chen B. P., Chen D. J., et al. (2005). Evidence for the direct binding of phosphorylated p53 to sites of DNA breaks in vivo. Cancer Res. 65, 10810–10821. - PubMed
-
- Arsham A. M., Howell J. J., Simon M. C. (2003). A novel hypoxia-inducible factor-independent hypoxic response regulating mammalian target of rapamycin and its targets. J. Biol. Chem. 278, 29655–29660. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
