

Tissue Distribution of Memory T and B Cells in Rhesus Monkeys following Influenza A Infection
- PMID: 26408671
- PMCID: PMC4642841
- DOI: 10.4049/jimmunol.1501702
Tissue Distribution of Memory T and B Cells in Rhesus Monkeys following Influenza A Infection
Abstract
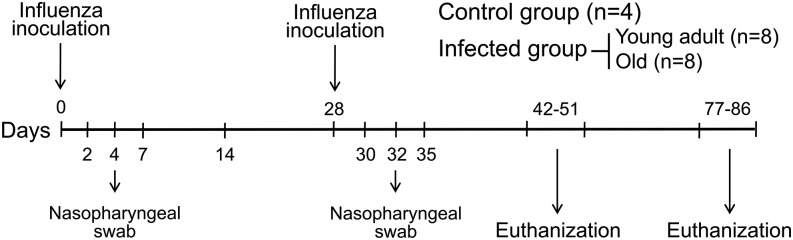
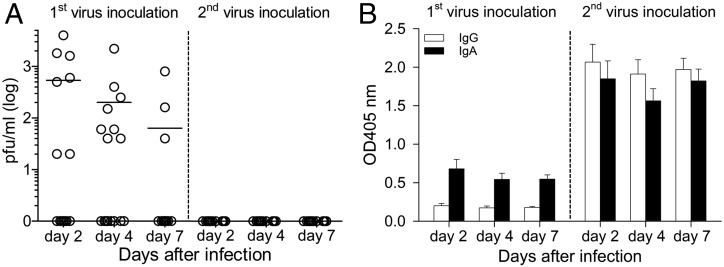
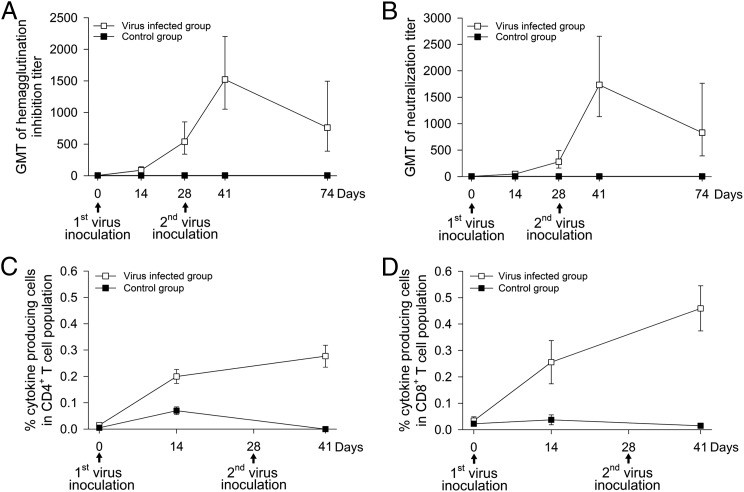
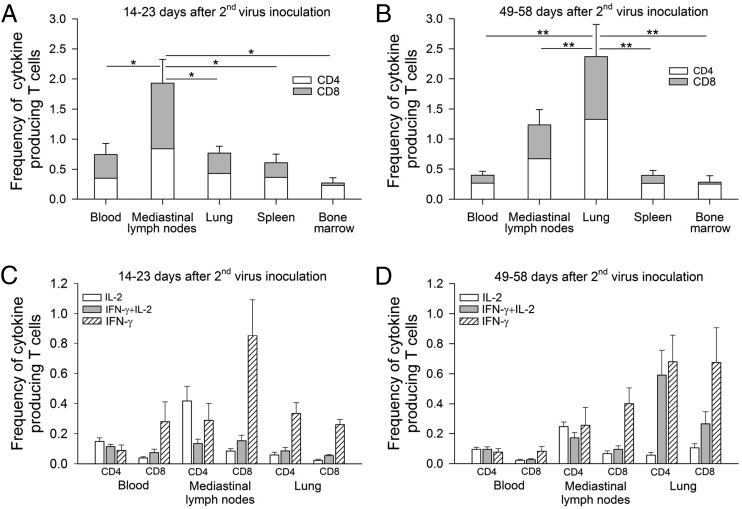
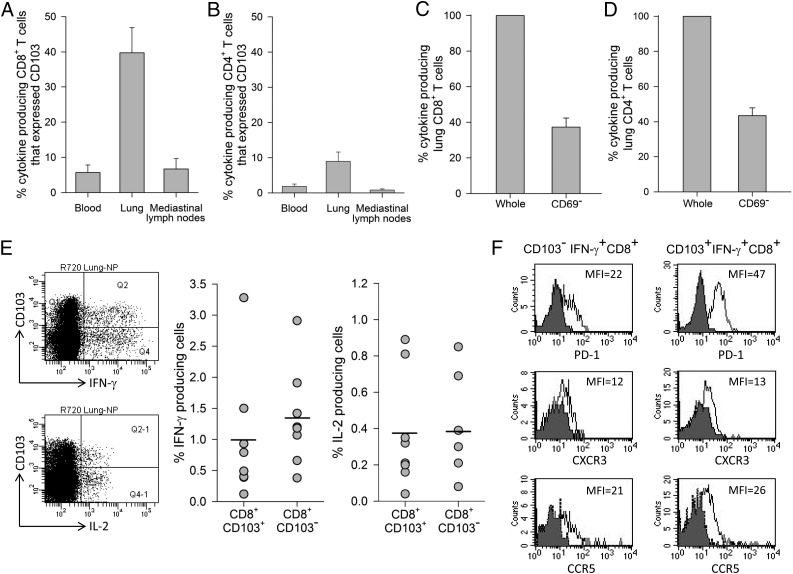
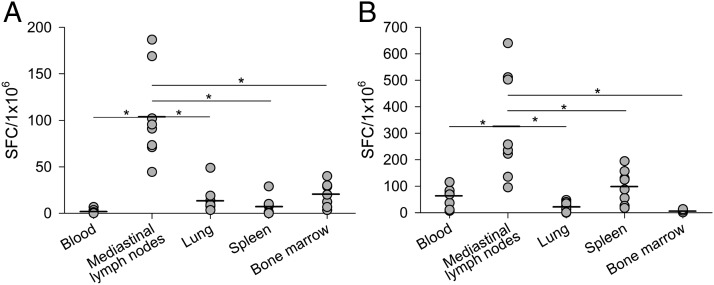
Studies of influenza-specific immune responses in humans have largely assessed systemic responses involving serum Ab and peripheral blood T cell responses. However, recent evidence indicates that tissue-resident memory T (TRM) cells play an important role in local murine intrapulmonary immunity. Rhesus monkeys were pulmonary exposed to 2009 pandemic H1N1 virus at days 0 and 28 and immune responses in different tissue compartments were measured. All animals were asymptomatic postinfection. Although only minimal memory immune responses were detected in peripheral blood, a high frequency of influenza nucleoprotein-specific memory T cells was detected in the lung at the "contraction phase," 49-58 d after second virus inoculation. A substantial proportion of lung nucleoprotein-specific memory CD8(+) T cells expressed CD103 and CD69, phenotypic markers of TRM cells. Lung CD103(+) and CD103(-) memory CD8(+) T cells expressed similar levels of IFN-γ and IL-2. Unlike memory T cells, spontaneous Ab secreting cells and memory B cells specific to influenza hemagglutinin were primarily observed in the mediastinal lymph nodes. Little difference in systemic and local immune responses against influenza was observed between young adult (6-8 y) and old animals (18-28 y). Using a nonhuman primate model, we revealed substantial induction of local T and B cell responses following 2009 pandemic H1N1 infection. Our study identified a subset of influenza-specific lung memory T cells characterized as TRM cells in rhesus monkeys. The rhesus monkey model may be useful to explore the role of TRM cells in local tissue protective immunity after rechallenge and vaccination.
Copyright © 2015 by The American Association of Immunologists, Inc.
Figures
References
-
- Osterholm M. T., Kelley N. S., Sommer A., Belongia E. A. 2012. Efficacy and effectiveness of influenza vaccines: a systematic review and meta-analysis. Lancet Infect. Dis. 12: 36–44. - PubMed
-
- Black S., Nicolay U., Vesikari T., Knuf M., Del Giudice G., Della Cioppa G., Tsai T., Clemens R., Rappuoli R. 2011. Hemagglutination inhibition antibody titers as a correlate of protection for inactivated influenza vaccines in children. Pediatr. Infect. Dis. J. 30: 1081–1085. - PubMed
-
- Gravenstein S., Drinka P., Duthie E. H., Miller B. A., Brown C. S., Hensley M., Circo R., Langer E., Ershler W. B. 1994. Efficacy of an influenza hemagglutinin-diphtheria toxoid conjugate vaccine in elderly nursing home subjects during an influenza outbreak. J. Am. Geriatr. Soc. 42: 245–251. - PubMed
-
- Yap K. L., Ada G. L. 1978. Cytotoxic T cells in the lungs of mice infected with an influenza A virus. Scand. J. Immunol. 7: 73–80. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
