

Drosophila spermatid individualization is sensitive to temperature and fatty acid metabolism
- PMID: 26413411
- PMCID: PMC4581069
- DOI: 10.1080/21565562.2015.1006089
Drosophila spermatid individualization is sensitive to temperature and fatty acid metabolism
Abstract
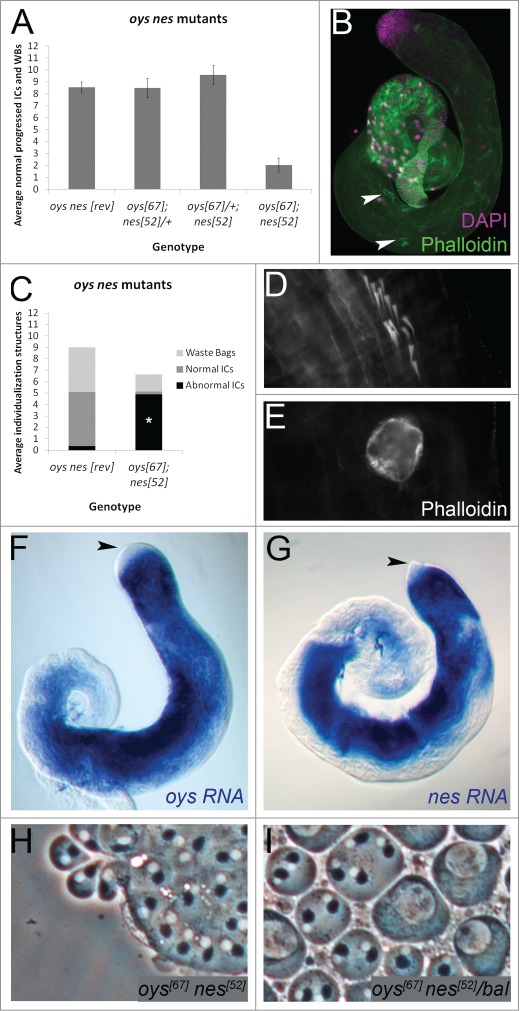
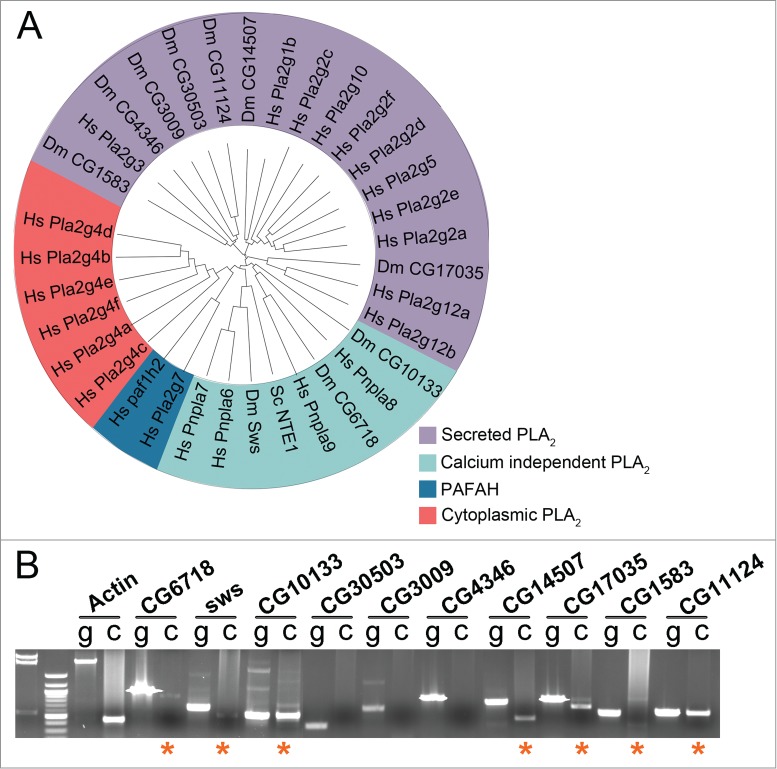
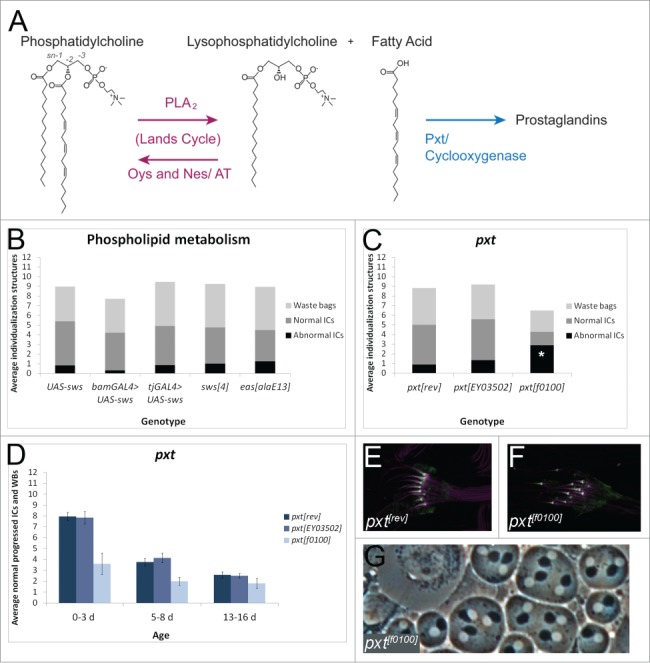
Fatty acids are precursors of potent lipid signaling molecules. They are stored in membrane phospholipids and released by phospholipase A2 (PLA2). Lysophospholipid acyltransferases (ATs) oppose PLA2 by re-esterifying fatty acids into phospholipids, in a biochemical pathway known as the Lands Cycle. Drosophila Lands Cycle ATs oys and nes, as well as 7 predicted PLA2 genes, are expressed in the male reproductive tract. Oys and Nes are required for spermatid individualization. Individualization, which occurs after terminal differentiation, invests each spermatid in its own plasma membrane and removes the bulk of the cytoplasmic contents. We developed a quantitative assay to measure individualization defects. We demonstrate that individualization is sensitive to temperature and age but not to diet. Mutation of the cyclooxygenase Pxt, which metabolizes fatty acids to prostaglandins, also leads to individualization defects. In contrast, modulating phospholipid levels by mutation of the phosphatidylcholine lipase Swiss cheese (Sws) or the ethanolamine kinase Easily shocked (Eas) does not perturb individualization, nor does Sws overexpression. Our results suggest that fatty acid derived signals such as prostaglandins, whose abundance is regulated by the Lands Cycle, are important regulators of spermatogenesis.
Keywords: actin; drosophila; fatty acids; individualization; lipid metabolism; prostaglandins; spermiogenesis.
Figures
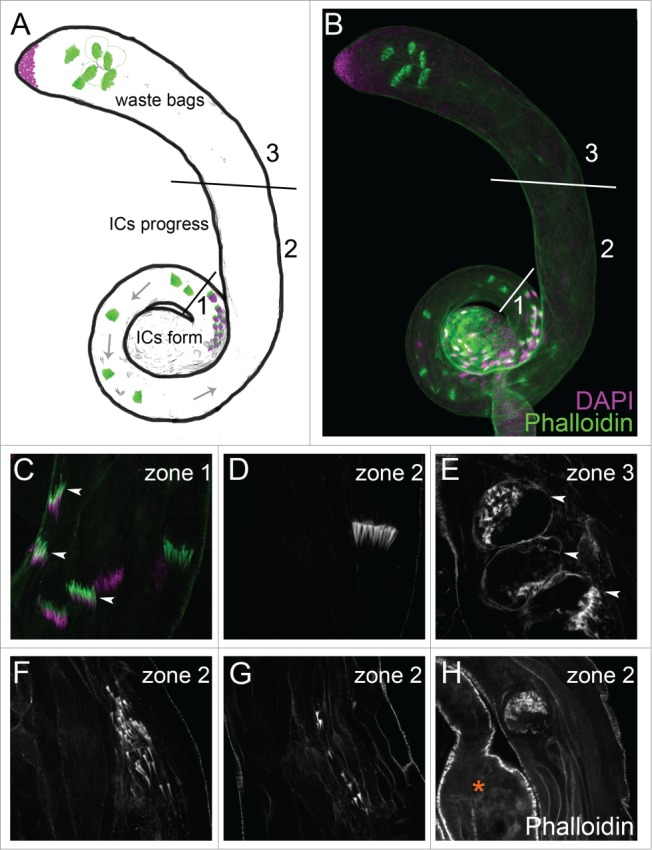
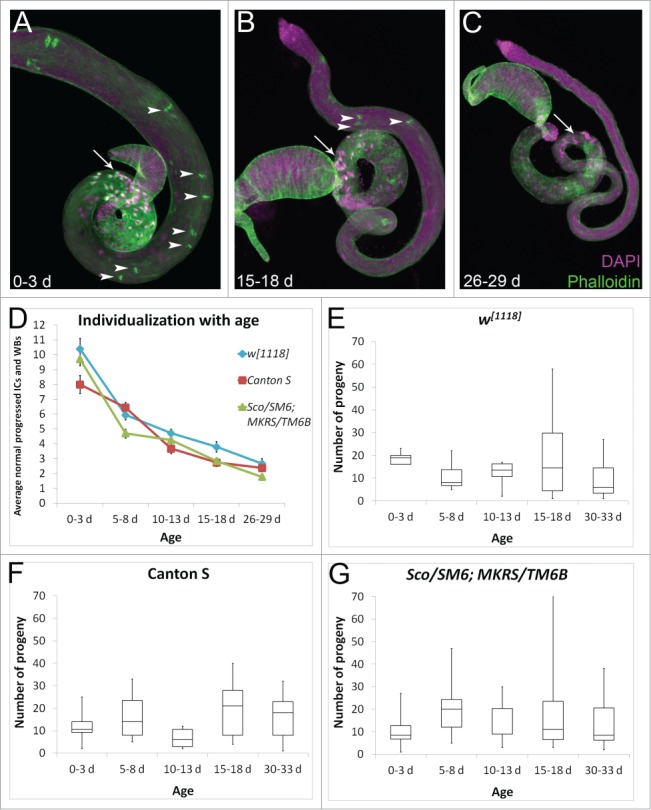
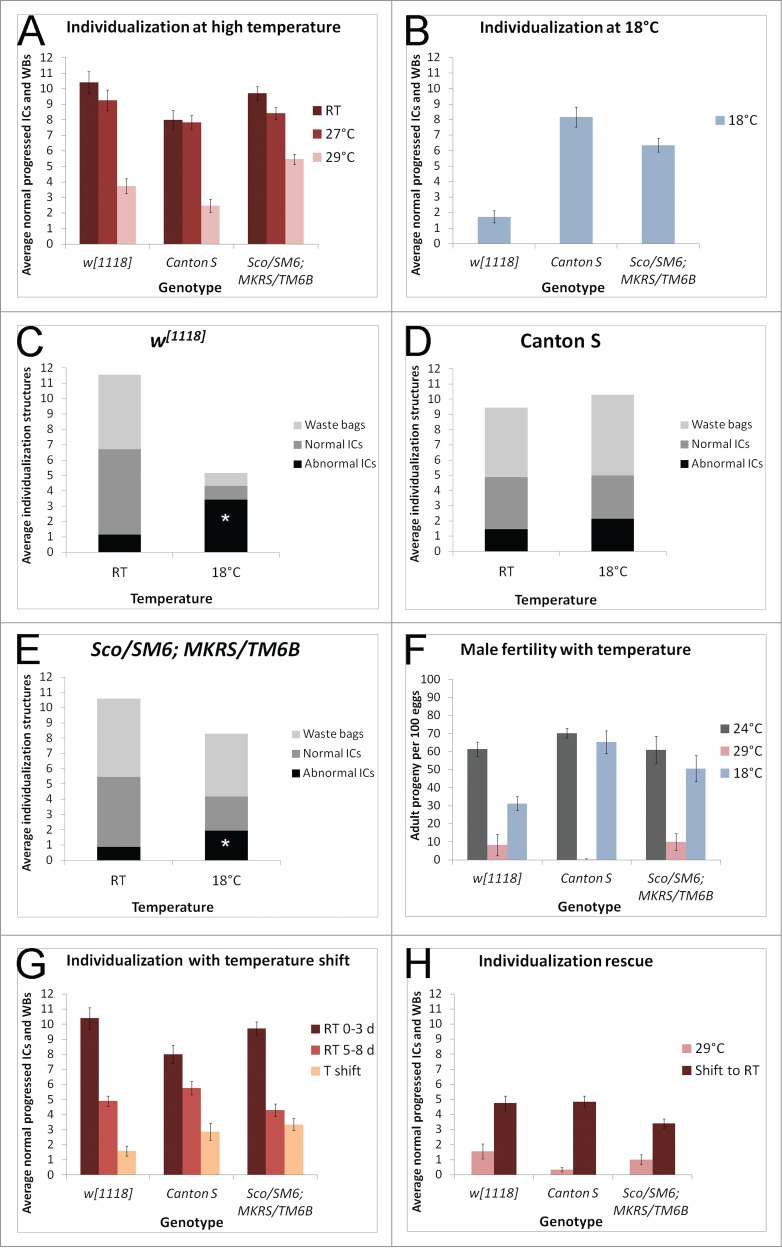
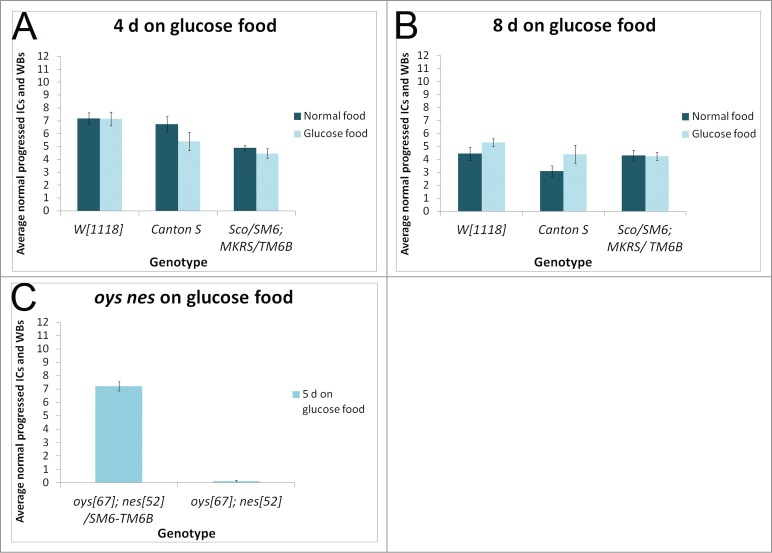
References
-
- Sutovsky P, Schatten G. Paternal contributions to the mammalian zygote: fertilization after sperm-egg fusion. Int Rev Cytol 2000; 195:1-65; PMID:10603574; http://dx.doi.org/10.1016/S0074-7696(08)62703-5 - DOI - PubMed
-
- Marcello MR, Singaravelu G, Singson A. Fertilization. Adv Exp Med Biol 2013; 757:321-50; PMID:22872482; http://dx.doi.org/10.1007/978-1-4614-4015-4_11 - DOI - PubMed
-
- Fitch KR, Yasuda GK, Owens KN, Wakimoto BT. Paternal effects in drosophila: implications for mechanisms of early development. Curr Topic Dev Biol 1998; 38:1-34; PMID:9399075; http://dx.doi.org/10.1016/S0070-2153(08)60243-4 - DOI - PubMed
-
- O'Donnell L, Nicholls PK, O'Bryan MK, McLachlan RI, Stanton PG. Spermiation: The process of sperm release. Spermatogenesis 2011; 1:14-35; PMID:21866274; http://dx.doi.org/10.4161/spmg.1.1.14525 - DOI - PMC - PubMed
-
- Fawcett DW, Ito S, Slautterback D. The occurrence of intercellular bridges in groups of cells exhibiting synchronous differentiation. J Biophysical Biochemical Cytol 1959; 5:453-60; PMID:13664686; http://dx.doi.org/10.1083/jcb.5.3.453 - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials