

Contribution of chaperones to STAT pathway signaling
- PMID: 26413421
- PMCID: PMC4580081
- DOI: 10.4161/21623988.2014.970459
Contribution of chaperones to STAT pathway signaling
Abstract
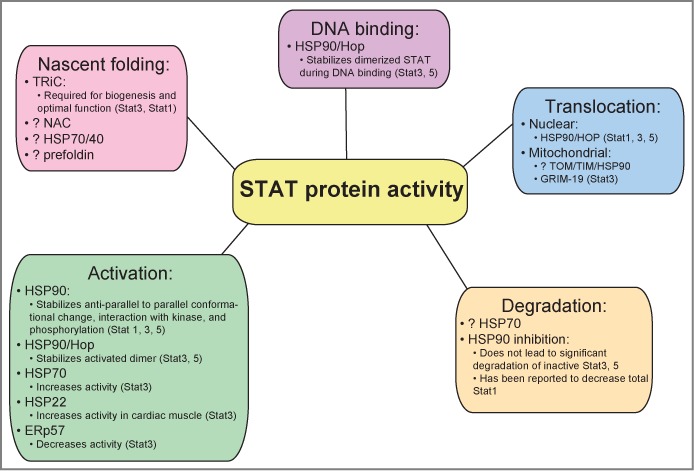
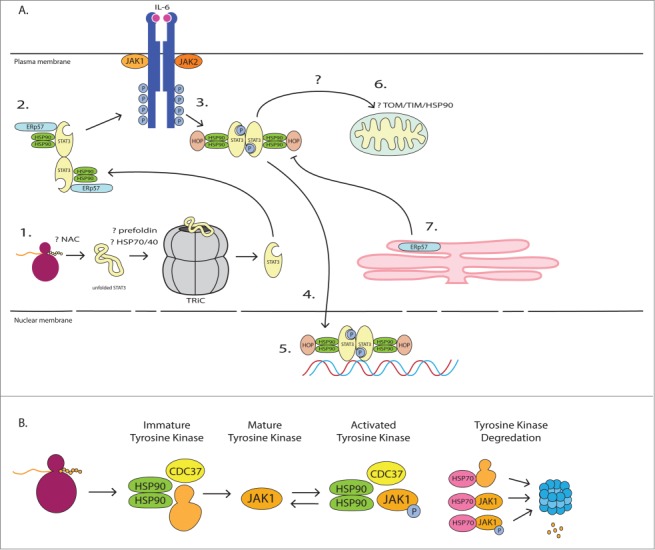
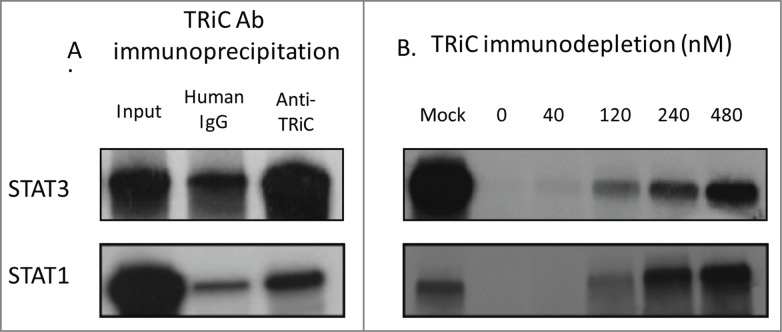
Aberrant STAT signaling is associated with the development and progression of many cancers and immune related diseases. Recent findings demonstrate that proteostasis modulators under clinical investigation for cancer therapy have a significant impact on STAT signaling, which may be critical for mediating their anti-cancer effects. Chaperones are critical for protein folding, stability and function and, thus, play an essential role in the maintenance of proteostasis. In this review we discuss the role of chaperones in STAT and tyrosine kinase (TK) protein folding, modulation of STAT and TK activity, and degradation of TKs. We highlight the important role of chaperones in STAT signaling, and how this knowledge has provided a framework for the development of new therapeutic avenues of targeting STAT signaling related pathologies.
Keywords: JAK; RTK; SRC; STAT; chaperone; chaperonin; heat shock proteins; tyrosine kinase.
Figures
References
-
- Powers ET, Morimoto RI, Dillin A, Kelly JW, Balch WE. Biological and chemical approaches to diseases of proteostasis deficiency. 2009; 78:959-91. Available from: www.annualreviews.orgdoiabs10.1146annurev.biochem.052308.114844">http:www.annualreviews.orgdoiabs10.1146annurev.biochem.052308.114844; PMID:19298183 - PubMed
-
- Hartl FU. Molecular chaperones in cellular protein folding. 1996; 381:571-9. Available from: http:eutils.ncbi.nlm.nih.goventrezeutilselink.fcgi?dbfrom=pubm-ed&id=863...; PMID:8637592 - PubMed
-
- Hartl FU, Hayer-Hartl M. Molecular chaperones in the cytosol: from nascent chain to folded protein. 2002; 295:1852-8. Available from: http:eutils.ncbi.nlm.nih.goventrezeutilselink.fcgi?dbfrom=pubmed&id=1188...; PMID:11884745 - PubMed
-
- Frydman J. Folding of newly translated proteins in vivo: the role of molecular chaperones. Annu Rev Biochem [Internet] 2001; 70:603-47. Available from: http:eutils.ncbi.nlm.nih.goventrezeutilselink.fcgi?dbfrom=pubmed&id=1139...; PMID:11395418; http://dx.doi.org/10.1146/annurev.biochem.70.1.603 - DOI - PubMed
-
- Bukau B, Horwich AL. The Hsp70 and Hsp60 chaperone machines. Cell 1998; 92:351-66; PMID:9476895; http://dx.doi.org/10.1016/S0092-8674(00)80928-9 - DOI - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous