

Multiple mechanisms for CRISPR-Cas inhibition by anti-CRISPR proteins
- PMID: 26416740
- PMCID: PMC4935067
- DOI: 10.1038/nature15254
Multiple mechanisms for CRISPR-Cas inhibition by anti-CRISPR proteins
Abstract
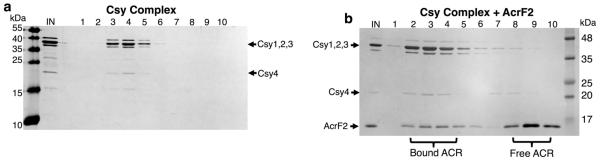
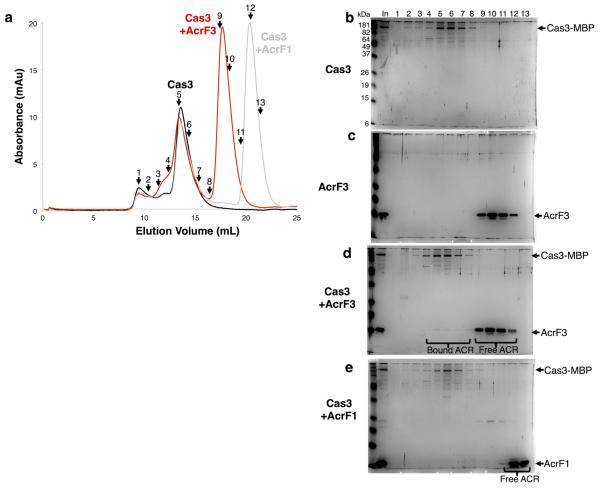
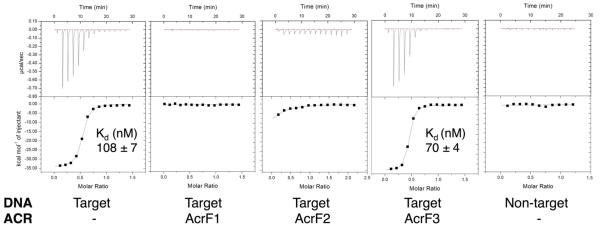
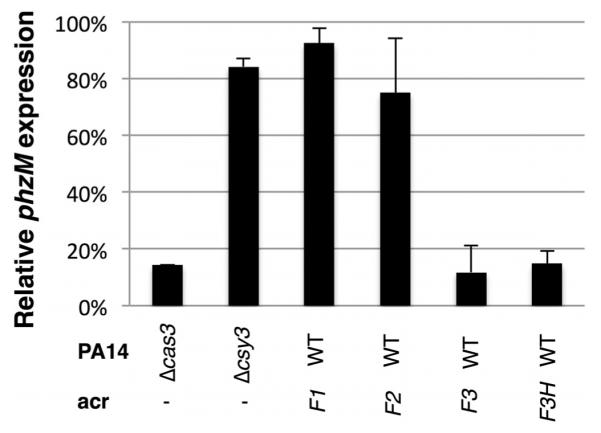
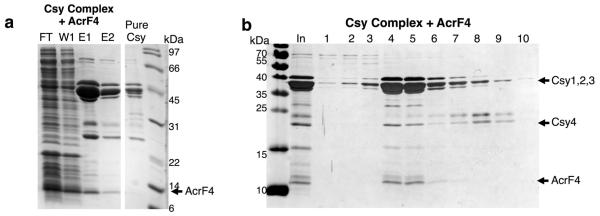
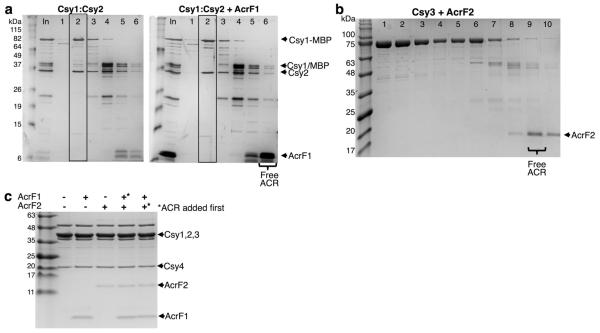
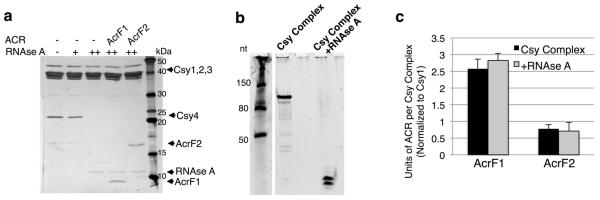
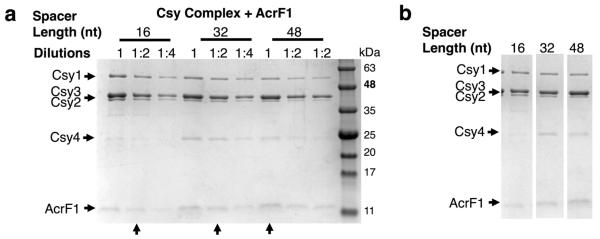
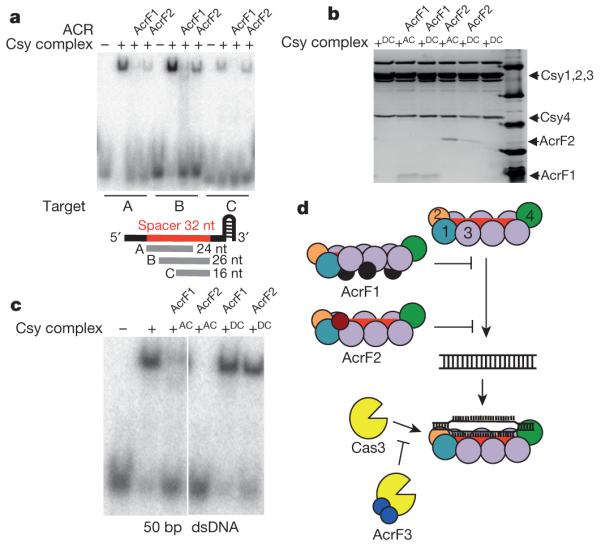
The battle for survival between bacteria and the viruses that infect them (phages) has led to the evolution of many bacterial defence systems and phage-encoded antagonists of these systems. Clustered regularly interspaced short palindromic repeats (CRISPR) and the CRISPR-associated (cas) genes comprise an adaptive immune system that is one of the most widespread means by which bacteria defend themselves against phages. We identified the first examples of proteins produced by phages that inhibit a CRISPR-Cas system. Here we performed biochemical and in vivo investigations of three of these anti-CRISPR proteins, and show that each inhibits CRISPR-Cas activity through a distinct mechanism. Two block the DNA-binding activity of the CRISPR-Cas complex, yet do this by interacting with different protein subunits, and using steric or non-steric modes of inhibition. The third anti-CRISPR protein operates by binding to the Cas3 helicase-nuclease and preventing its recruitment to the DNA-bound CRISPR-Cas complex. In vivo, this anti-CRISPR can convert the CRISPR-Cas system into a transcriptional repressor, providing the first example-to our knowledge-of modulation of CRISPR-Cas activity by a protein interactor. The diverse sequences and mechanisms of action of these anti-CRISPR proteins imply an independent evolution, and foreshadow the existence of other means by which proteins may alter CRISPR-Cas function.
Figures
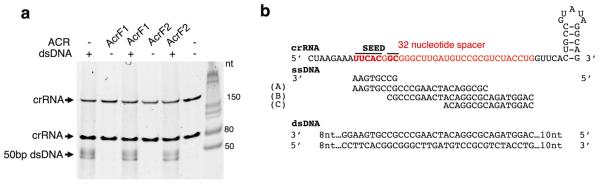
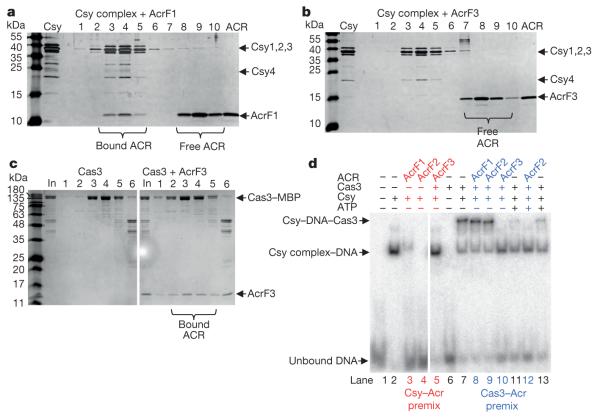
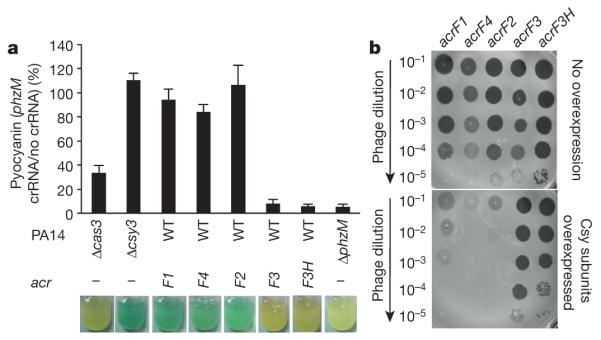
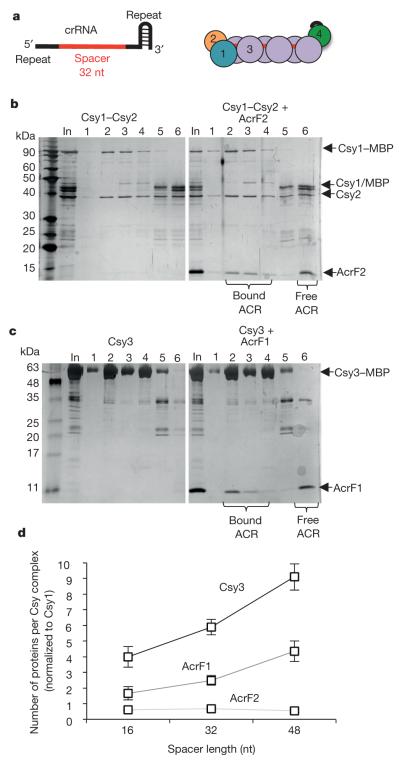
Comment in
-
Microbial genetics: Phages' box of tricks for CRISPR.Nat Rev Genet. 2015 Nov;16(11):626. doi: 10.1038/nrg4030. Epub 2015 Oct 13. Nat Rev Genet. 2015. PMID: 26460346 No abstract available.
References
-
- Barrangou R, et al. CRISPR provides acquired resistance against viruses in prokaryotes. Science. 2007;315:1709–1712. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
