

Regulation of calreticulin-major histocompatibility complex (MHC) class I interactions by ATP
- PMID: 26420867
- PMCID: PMC4611598
- DOI: 10.1073/pnas.1510132112
Regulation of calreticulin-major histocompatibility complex (MHC) class I interactions by ATP
Abstract
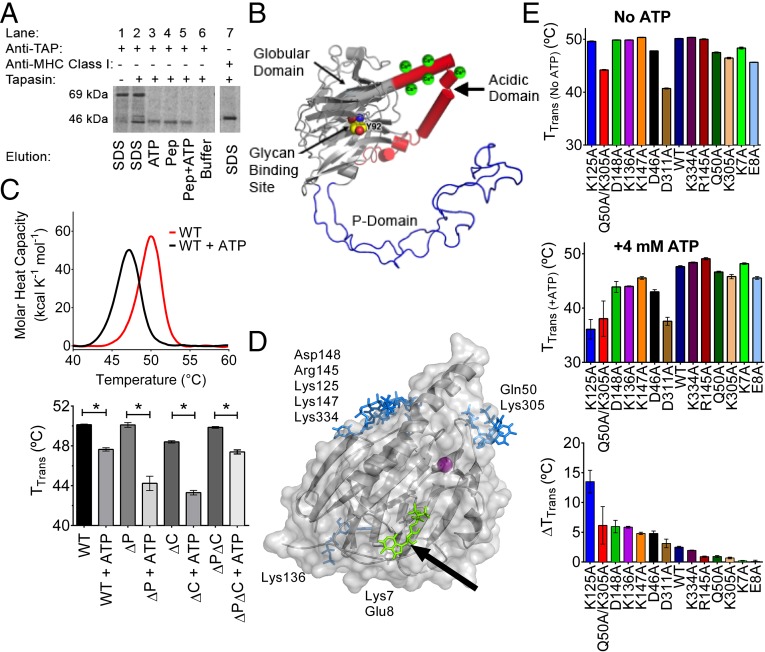
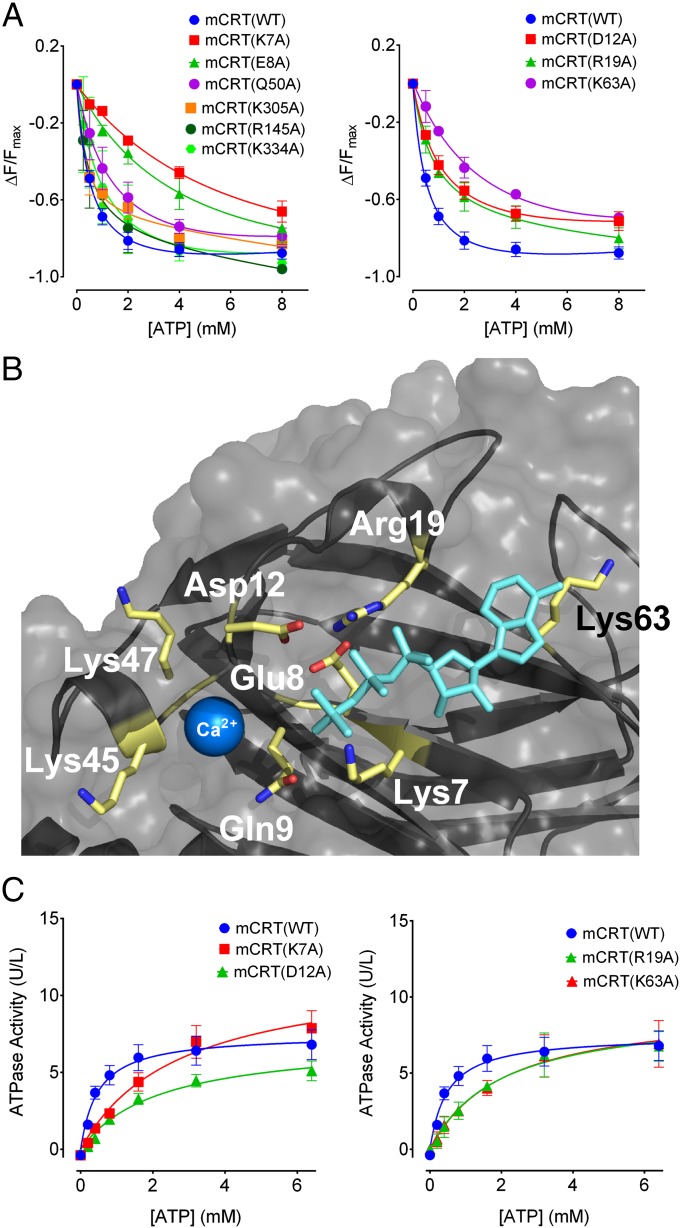
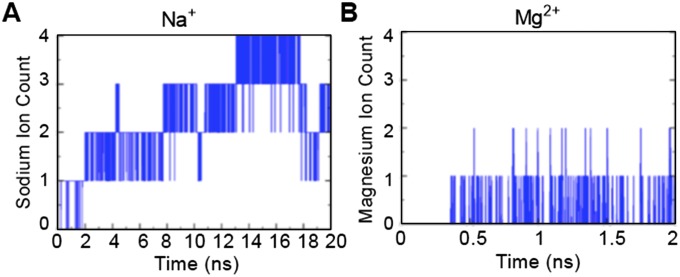
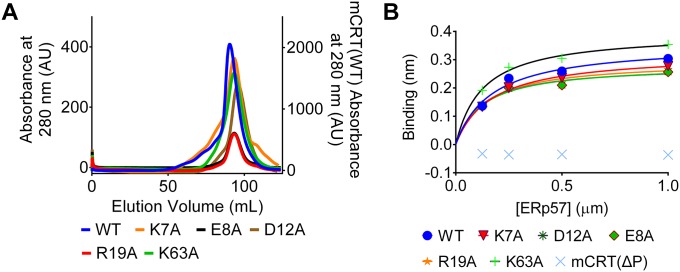
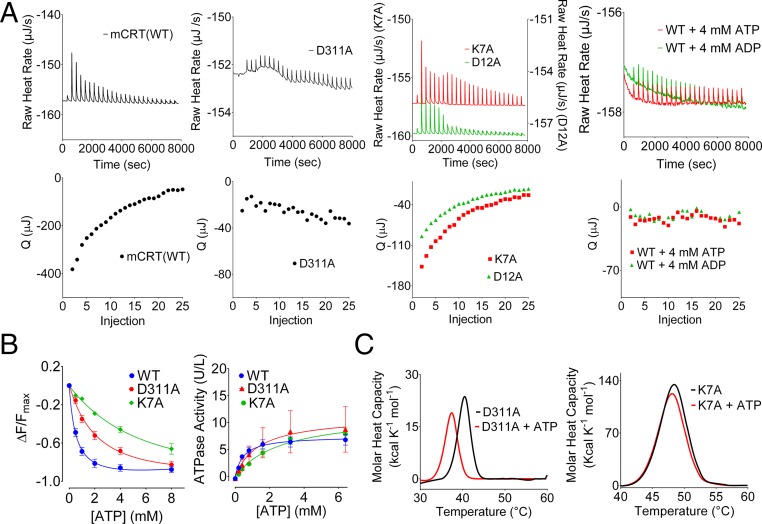
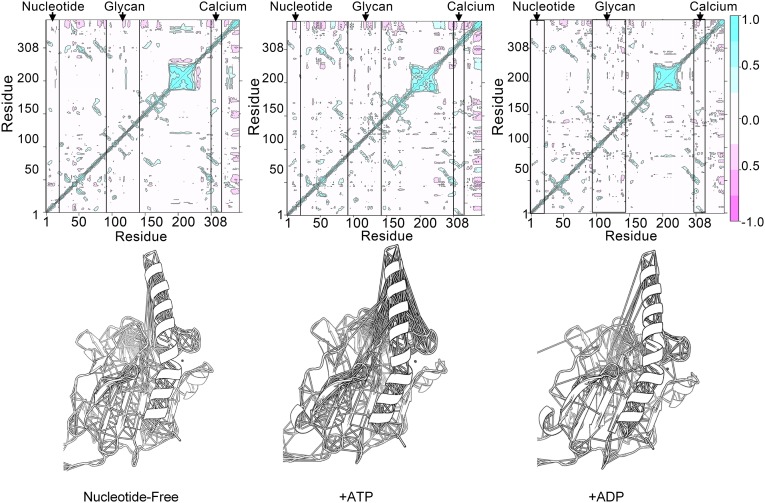
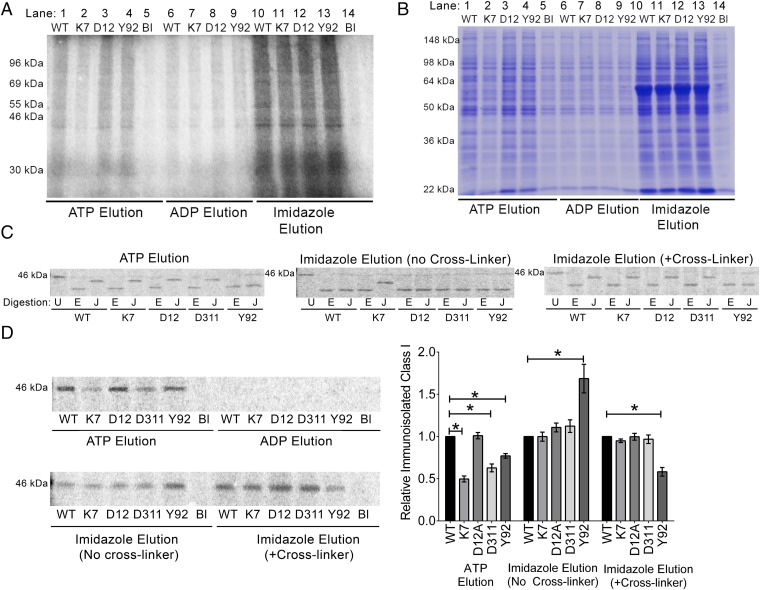
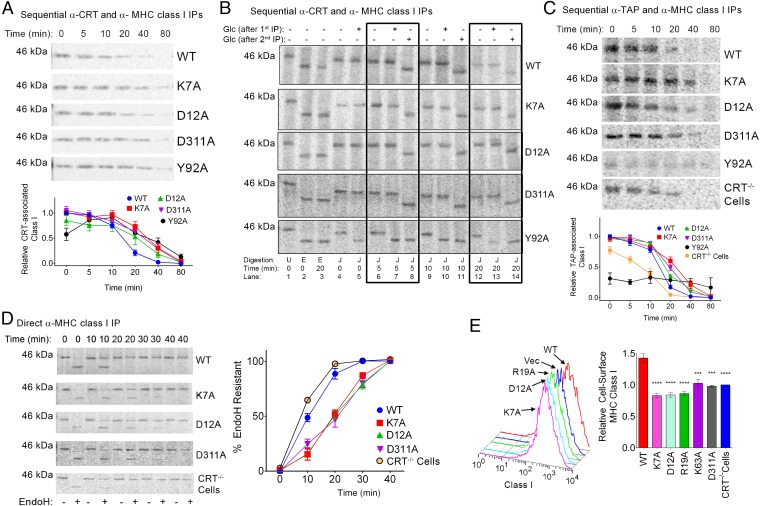
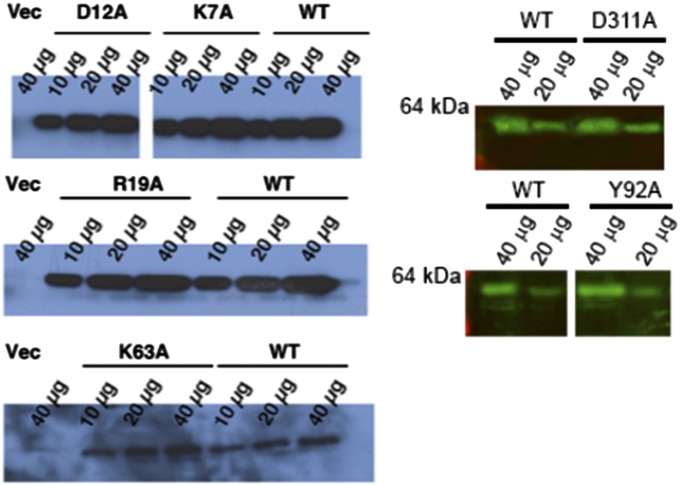
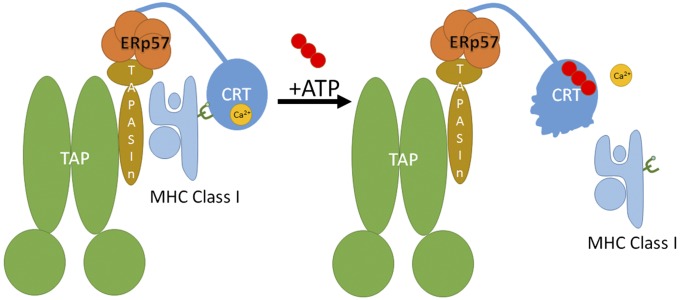
The MHC class I peptide loading complex (PLC) facilitates the assembly of MHC class I molecules with peptides, but factors that regulate the stability and dynamics of the assembly complex are largely uncharacterized. Based on initial findings that ATP, in addition to MHC class I-specific peptide, is able to induce MHC class I dissociation from the PLC, we investigated the interaction of ATP with the chaperone calreticulin, an endoplasmic reticulum (ER) luminal, calcium-binding component of the PLC that is known to bind ATP. We combined computational and experimental measurements to identify residues within the globular domain of calreticulin, in proximity to the high-affinity calcium-binding site, that are important for high-affinity ATP binding and for ATPase activity. High-affinity calcium binding by calreticulin is required for optimal nucleotide binding, but both ATP and ADP destabilize enthalpy-driven high-affinity calcium binding to calreticulin. ATP also selectively destabilizes the interaction of calreticulin with cellular substrates, including MHC class I molecules. Calreticulin mutants that affect ATP or high-affinity calcium binding display prolonged associations with monoglucosylated forms of cellular MHC class I, delaying MHC class I dissociation from the PLC and their transit through the secretory pathway. These studies reveal central roles for ATP and calcium binding as regulators of calreticulin-substrate interactions and as key determinants of PLC dynamics.
Keywords: ATP; ATPase; MHC class; calreticulin; monoglucosylated glycans.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Gao B, et al. Assembly and antigen-presenting function of MHC class I molecules in cells lacking the ER chaperone calreticulin. Immunity. 2002;16(1):99–109. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
