

Conserved Motifs within Hepatitis C Virus Envelope (E2) RNA and Protein Independently Inhibit T Cell Activation
- PMID: 26421924
- PMCID: PMC4589396
- DOI: 10.1371/journal.ppat.1005183
Conserved Motifs within Hepatitis C Virus Envelope (E2) RNA and Protein Independently Inhibit T Cell Activation
Abstract
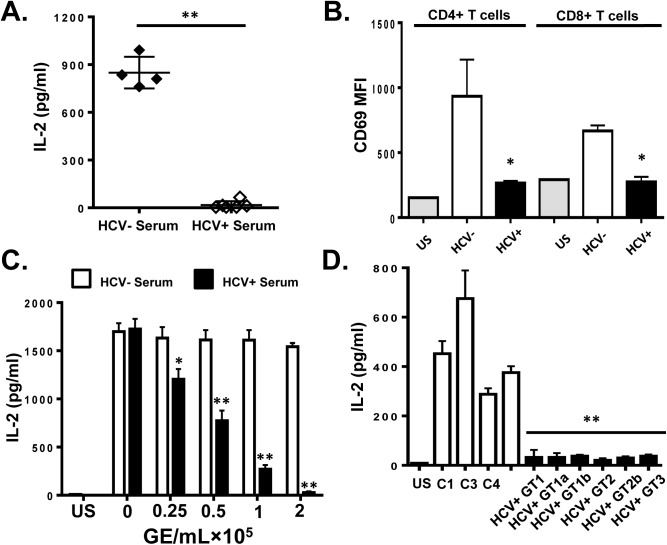
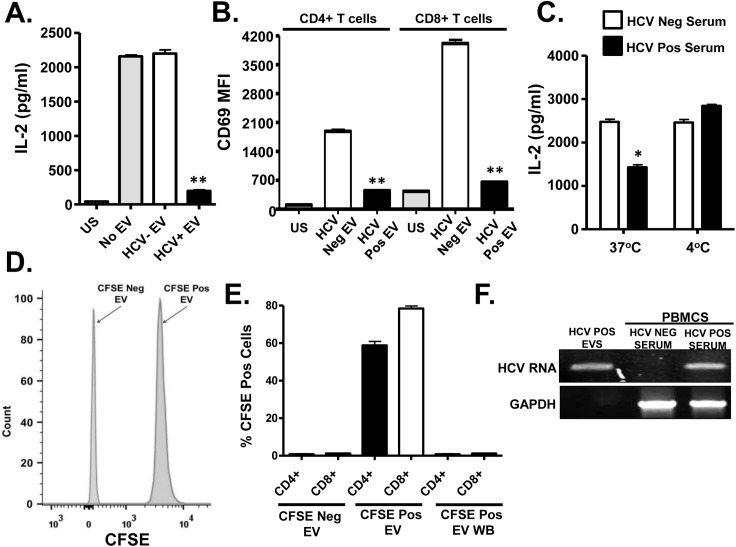
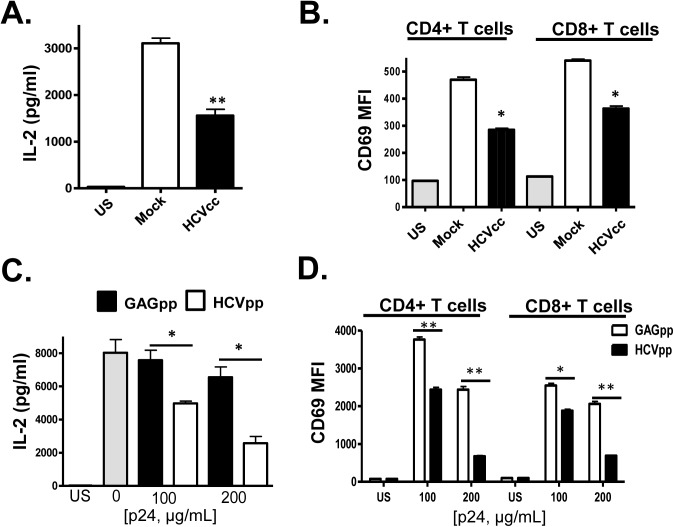
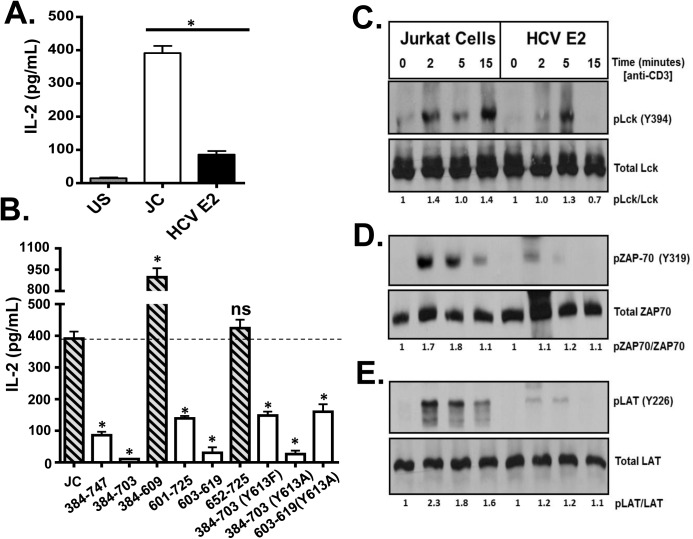
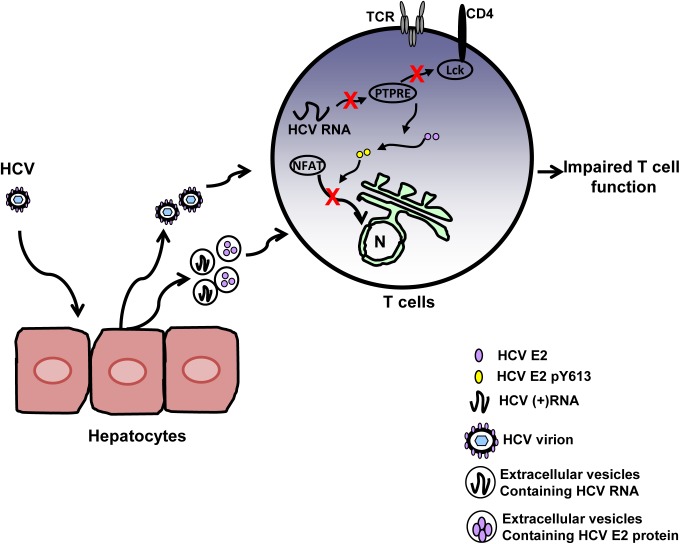
T cell receptor (TCR) signaling is required for T-cell activation, proliferation, differentiation, and effector function. Hepatitis C virus (HCV) infection is associated with impaired T-cell function leading to persistent viremia, delayed and inconsistent antibody responses, and mild immune dysfunction. Although multiple factors appear to contribute to T-cell dysfunction, a role for HCV particles in this process has not been identified. Here, we show that incubation of primary human CD4+ and CD8+ T-cells with HCV RNA-containing serum, HCV-RNA containing extracellular vesicles (EVs), cell culture derived HCV particles (HCVcc) and HCV envelope pseudotyped retrovirus particles (HCVpp) inhibited TCR-mediated signaling. Since HCVpp's contain only E1 and E2, we examined the effect of HCV E2 on TCR signaling pathways. HCV E2 expression recapitulated HCV particle-induced TCR inhibition. A highly conserved, 51 nucleotide (nt) RNA sequence was sufficient to inhibit TCR signaling. Cells expressing the HCV E2 coding RNA contained a short, virus-derived RNA predicted to be a Dicer substrate, which targeted a phosphatase involved in Src-kinase signaling (PTPRE). T-cells and hepatocytes containing HCV E2 RNA had reduced PTPRE protein levels. Mutation of 6 nts abolished the predicted Dicer interactions and restored PTPRE expression and proximal TCR signaling. HCV RNA did not inhibit distal TCR signaling induced by PMA and Ionomycin; however, HCV E2 protein inhibited distal TCR signaling. This inhibition required lymphocyte-specific tyrosine kinase (Lck). Lck phosphorylated HCV E2 at a conserved tyrosine (Y613), and phospho-E2 inhibited nuclear translocation of NFAT. Mutation of Y613 restored distal TCR signaling, even in the context of HCVpps. Thus, HCV particles delivered viral RNA and E2 protein to T-cells, and these inhibited proximal and distal TCR signaling respectively. These effects of HCV particles likely aid in establishing infection and contribute to viral persistence.
Conflict of interest statement
I have read the journal's policy and the authors of this manuscript have the following competing interests: Drs. Bhattarai, McLinden, Xiang, and Stapleton have patents or provisional patents related to the use of HCV E2 RNA or protein for therapeutic purposes. This does not alter our adherence to all PLOS policies on sharing data and materials.
Figures
Similar articles
-
Hepatitis C virus infection inhibits a Src-kinase regulatory phosphatase and reduces T cell activation in vivo.PLoS Pathog. 2017 Feb 24;13(2):e1006232. doi: 10.1371/journal.ppat.1006232. eCollection 2017 Feb. PLoS Pathog. 2017. PMID: 28235043 Free PMC article.
-
GB virus C particles inhibit T cell activation via envelope E2 protein-mediated inhibition of TCR signaling.J Immunol. 2013 Jun 15;190(12):6351-9. doi: 10.4049/jimmunol.1300589. Epub 2013 May 17. J Immunol. 2013. PMID: 23686495 Free PMC article.
-
A novel T cell evasion mechanism in persistent RNA virus infection.Trans Am Clin Climatol Assoc. 2014;125:14-24; discussion 24-6. Trans Am Clin Climatol Assoc. 2014. PMID: 25125715 Free PMC article. Review.
-
Chimeric antigen receptor (CAR)-engineered T cells redirected against hepatitis C virus (HCV) E2 glycoprotein.Gut. 2016 Mar;65(3):512-23. doi: 10.1136/gutjnl-2014-308316. Epub 2015 Feb 6. Gut. 2016. PMID: 25661083 Free PMC article.
-
T cell recognition of hepatitis B and C viral antigens.Eur J Clin Invest. 1994 Oct;24(10):641-50. doi: 10.1111/j.1365-2362.1994.tb01055.x. Eur J Clin Invest. 1994. PMID: 7531642 Review.
Cited by
-
Extracellular vesicles and viruses: Are they close relatives?Proc Natl Acad Sci U S A. 2016 Aug 16;113(33):9155-61. doi: 10.1073/pnas.1605146113. Epub 2016 Jul 18. Proc Natl Acad Sci U S A. 2016. PMID: 27432966 Free PMC article.
-
From Structural Studies to HCV Vaccine Design.Viruses. 2021 May 4;13(5):833. doi: 10.3390/v13050833. Viruses. 2021. PMID: 34064532 Free PMC article. Review.
-
Yellow Fever Virus, but Not Zika Virus or Dengue Virus, Inhibits T-Cell Receptor-Mediated T-Cell Function by an RNA-Based Mechanism.J Infect Dis. 2017 Nov 27;216(9):1164-1175. doi: 10.1093/infdis/jix462. J Infect Dis. 2017. PMID: 28968905 Free PMC article.
-
Occult Infection with Hepatitis C Virus: Looking for Clear-Cut Boundaries and Methodological Consensus.J Clin Med. 2021 Dec 14;10(24):5874. doi: 10.3390/jcm10245874. J Clin Med. 2021. PMID: 34945170 Free PMC article. Review.
-
Vehicles of intercellular communication: exosomes and HIV-1.J Gen Virol. 2019 Mar;100(3):350-366. doi: 10.1099/jgv.0.001193. Epub 2019 Jan 31. J Gen Virol. 2019. PMID: 30702421 Free PMC article. Review.
References
-
- Wedemeyer H, He XS, Nascimbeni M, Davis AR, Greenberg HB, et al. (2002) Impaired effector function of hepatitis C virus-specific CD8+ T cells in chronic hepatitis C virus infection. J Immunol 169: 3447–3458. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
