

B1-Metallo-β-Lactamases: Where Do We Stand?
- PMID: 26424398
- PMCID: PMC4814356
- DOI: 10.2174/1389450116666151001105622
B1-Metallo-β-Lactamases: Where Do We Stand?
Abstract
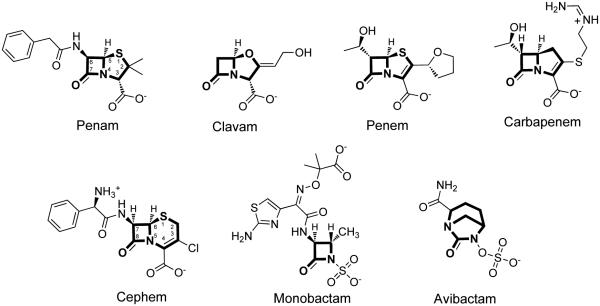
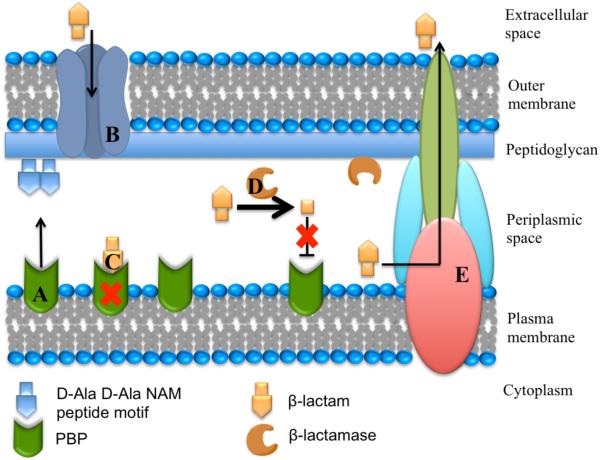
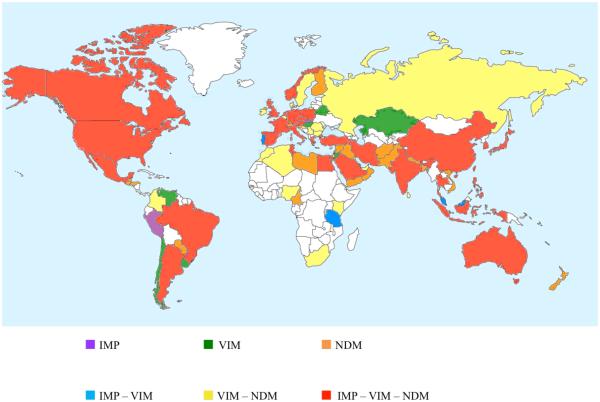
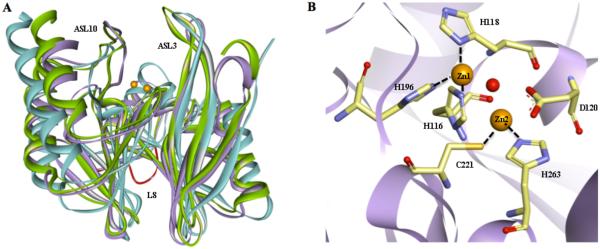
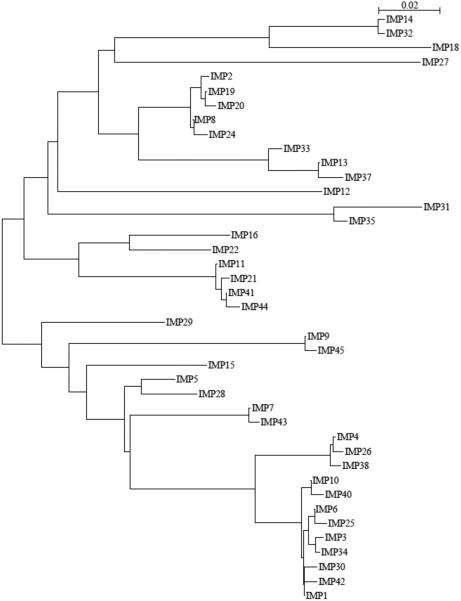
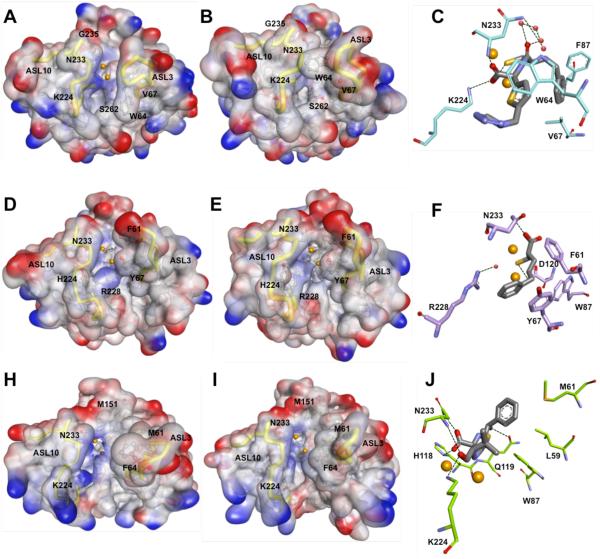
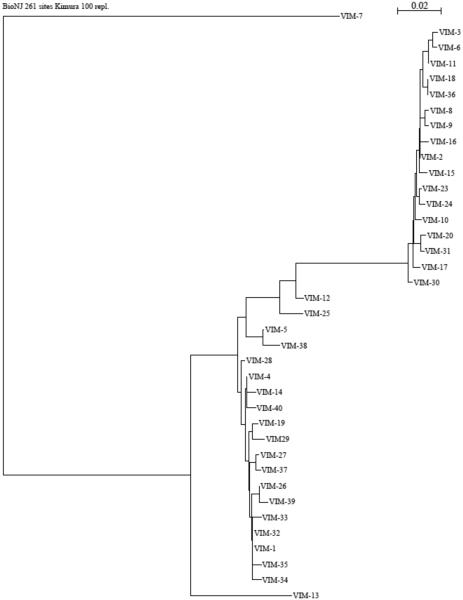
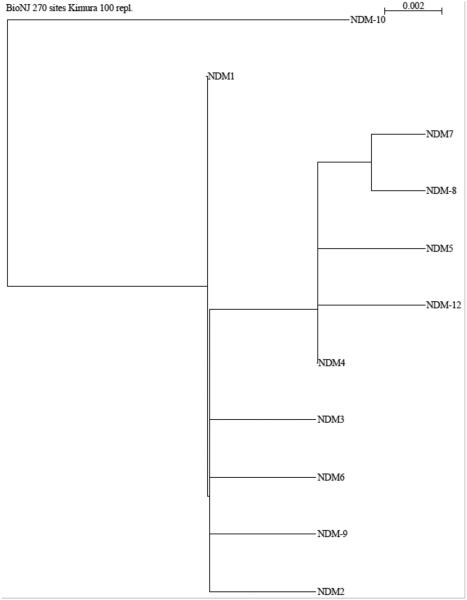
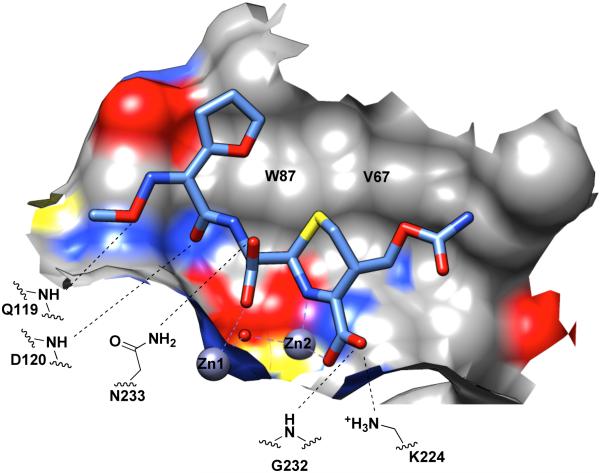
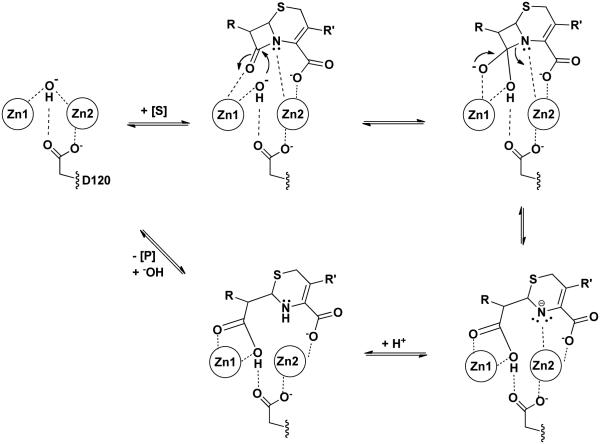
Metallo-β-Lactamases (MBLs) are class Bβ-lactamases that hydrolyze almost all clinically-availableβ-lactam antibiotics. MBLs feature the distinctive αβ/βα sandwich fold of the metallo-hydrolase/oxidoreductase superfamily and possess a shallow active-site groove containing one or two divalent zinc ions, flanked by flexible loops. According to sequence identity and zinc ion dependence, MBLs are classified into three subclasses (B1, B2 and B3), of which the B1 subclass enzymes have emerged as the most clinically significant. Differences among the active site architectures, the nature of zinc ligands, and the catalytic mechanisms have limited the development of a common inhibitor. In this review, we will describe the molecular epidemiology and structural studies of the most prominent representatives of class B1 MBLs (NDM-1, IMP-1 and VIM-2) and describe the implications for inhibitor design to counter this growing clinical threat.
Figures

References
-
- Fleming A. Classics in infectious diseases: on the antibacterial action of cultures of a penicillium, with special reference to their use in the isolation of B. influenzae by Alexander Fleming, Reprinted from the British Journal of Experimental Pathology 10:226-236, 1929. Rev Infect Dis. 1980;2:129–39. - PubMed
-
- Waxman DJ, Strominger JL. Penicillin-binding proteins and the mechanism of action of beta-lactam antibiotics. Annu Rev Biochem. 1983;52:825–69. - PubMed
-
- Page MI. The Mechanisms of Reactions of Beta-Lactam Antibiotics. Adv Phys Org Chem. 1987;23:165–270.
-
- Frere JM, Joris B. Penicillin-sensitive enzymes in peptidoglycan biosynthesis. Crit Rev Microbiol. 1985;11:299–396. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous