

Acetylation of NDPK-D Regulates Its Subcellular Localization and Cell Survival
- PMID: 26426123
- PMCID: PMC4591271
- DOI: 10.1371/journal.pone.0139616
Acetylation of NDPK-D Regulates Its Subcellular Localization and Cell Survival
Abstract
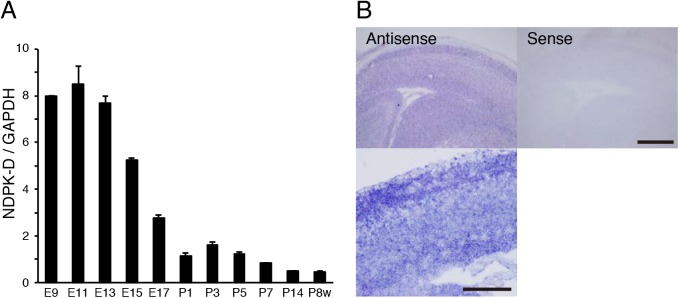
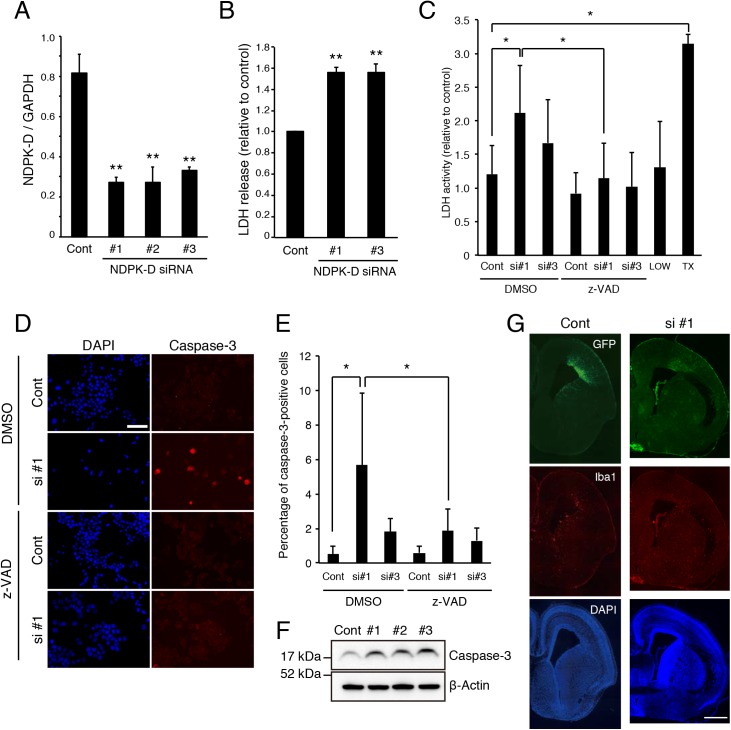
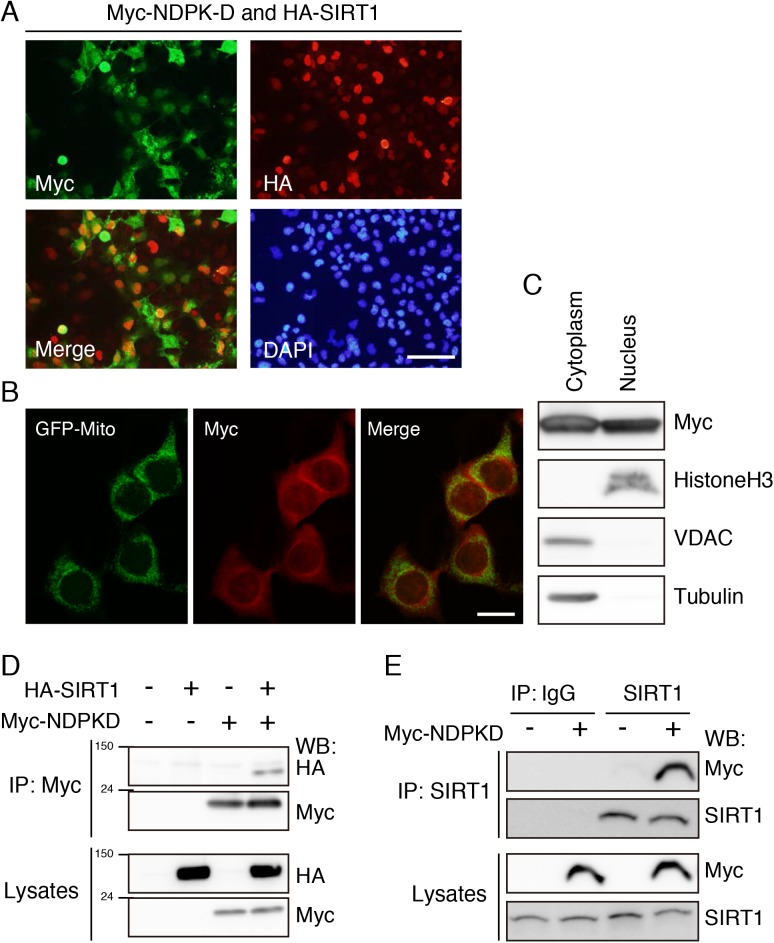
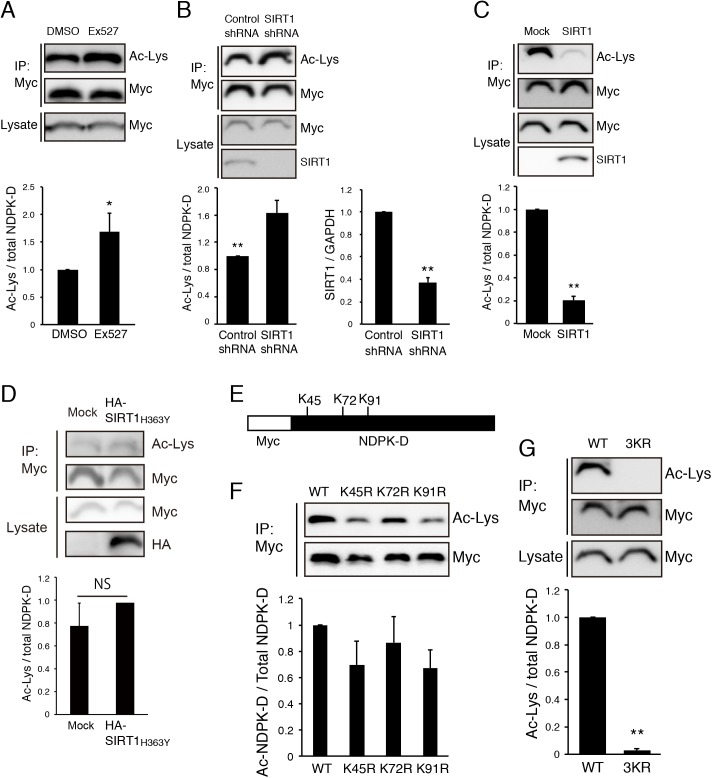
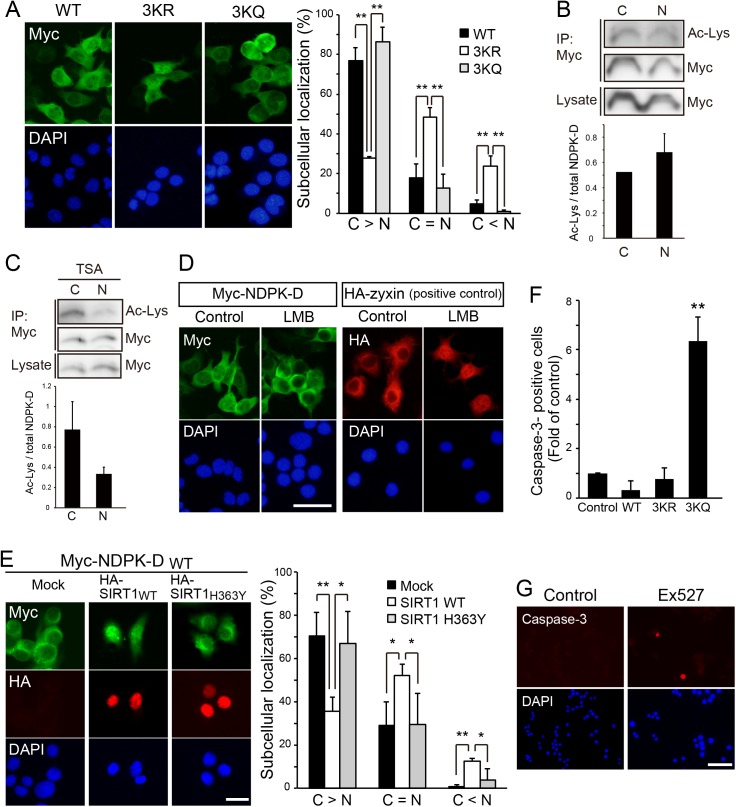
Nucleoside diphosphate kinases (NDPK) are ubiquitous enzymes that catalyze the reversible phosphotransfer of γ-phosphates between di- and triphosphonucleosides. NDPK-D (Nm23-H4) is the only member of the NDPK family with a mitochondrial targeting sequence. Despite the high expression of NDPK-D in the developing central nervous system, its function remains to be determined. In this study, we show that NDPK-D knockdown induces apoptosis in neuroblastoma cells as well as in mouse cortex, suggesting that NDPK-D is required for neuronal survival. We identified NDPK-D as a binding partner of NAD+-dependent histone deacetylase, SIRT1, by yeast two-hybrid screening. NDPK-D co-localized with SIRT1, and the association of these molecules was confirmed by co-immunoprecipitation. Inhibition of SIRT1 increases the acetylation of NDPK-D. Overexpression of NDPK-D along with SIRT1, or mutation in the acetylated lysine residues in NDPK-D, increases its nuclear accumulation. Furthermore, the NDPK-D acetylation-mimic mutant increased apoptosis in N1E-115 cells. Our data demonstrate that acetylation regulates the shuttling of NDPK-D between nucleus and cytoplasm, and increased acetylation of NDPK-D causes apoptosis.
Conflict of interest statement
Figures
References
-
- Frye RA. Characterization of five human cDNAs with homology to the yeast SIR2 gene: Sir2-like proteins (sirtuins) metabolize NAD and may have protein ADP-ribosyltransferase activity. Biochem Biophys Res Commun. 1999;260(1): 273–9. - PubMed
-
- Tanny JC, Dowd GJ, Huang J, Hilz H, Moazed D. An enzymatic activity in the yeast Sir2 protein that is essential for gene silencing. Cell. 1999;99(7): 735–45. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
