

Disruption of Sphingolipid Biosynthesis Blocks Phagocytosis of Candida albicans
- PMID: 26431038
- PMCID: PMC4592247
- DOI: 10.1371/journal.ppat.1005188
Disruption of Sphingolipid Biosynthesis Blocks Phagocytosis of Candida albicans
Abstract
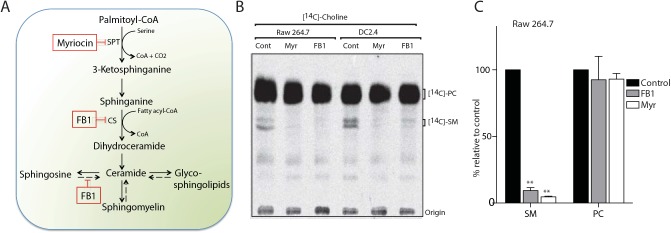
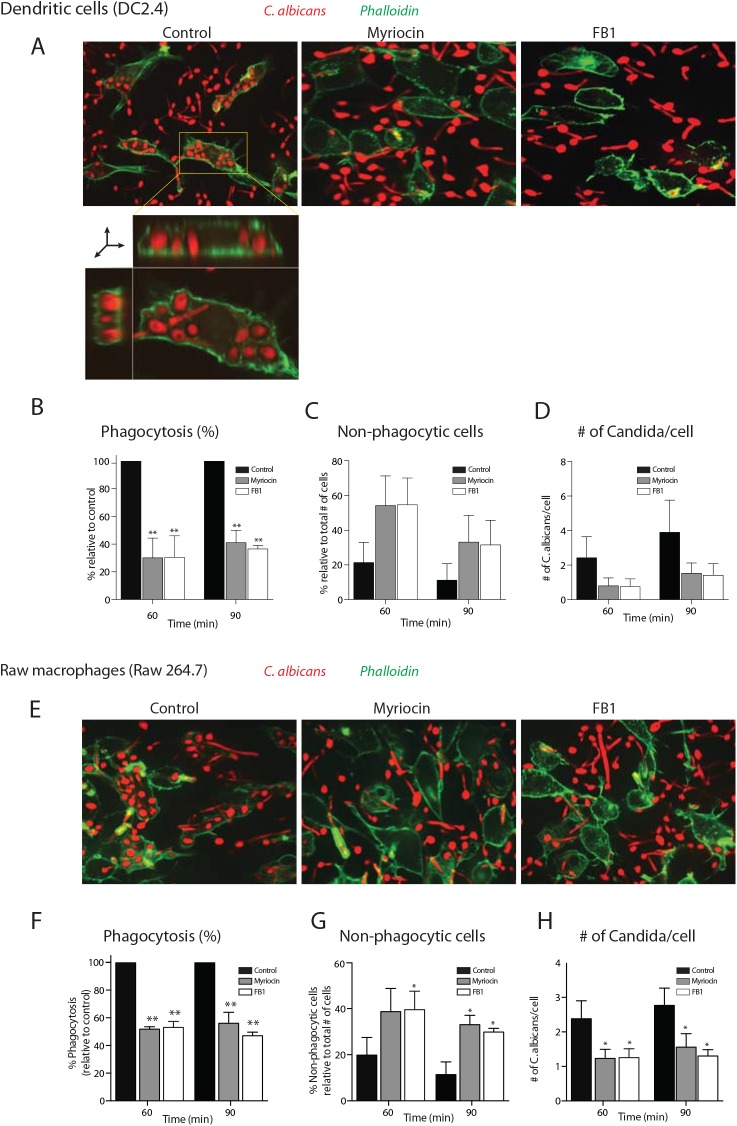
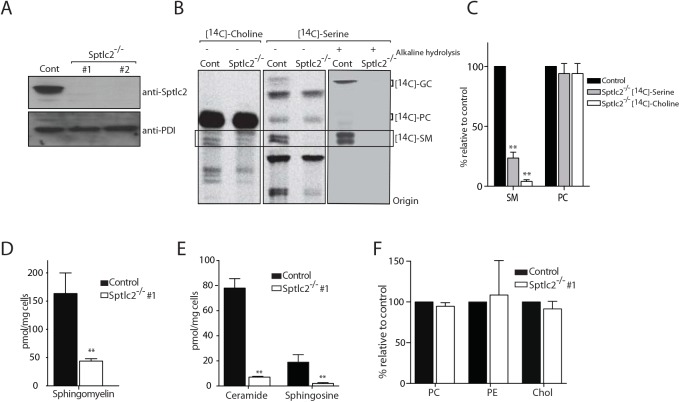
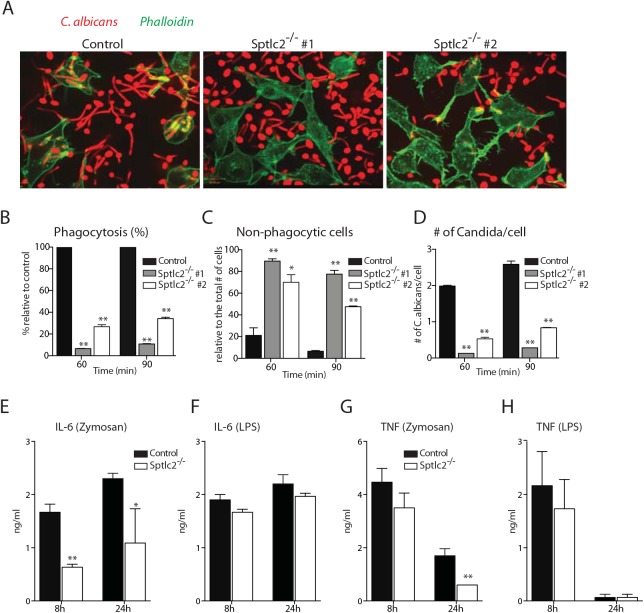
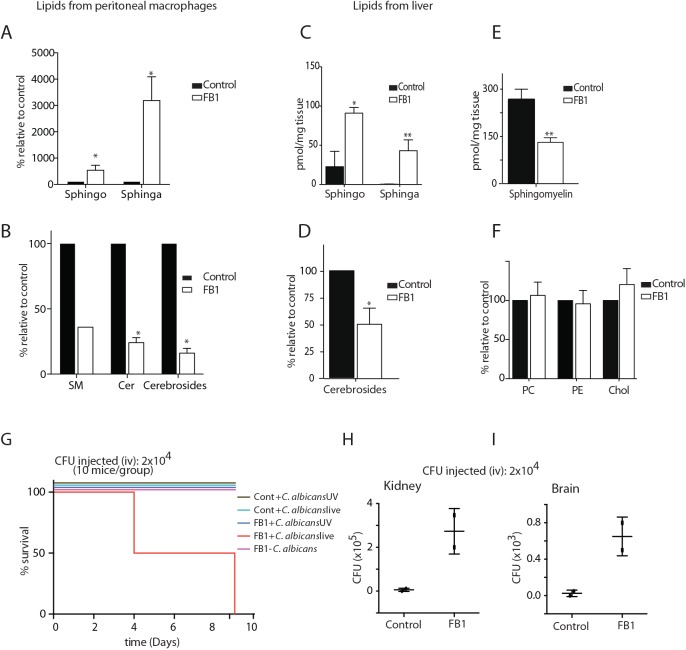
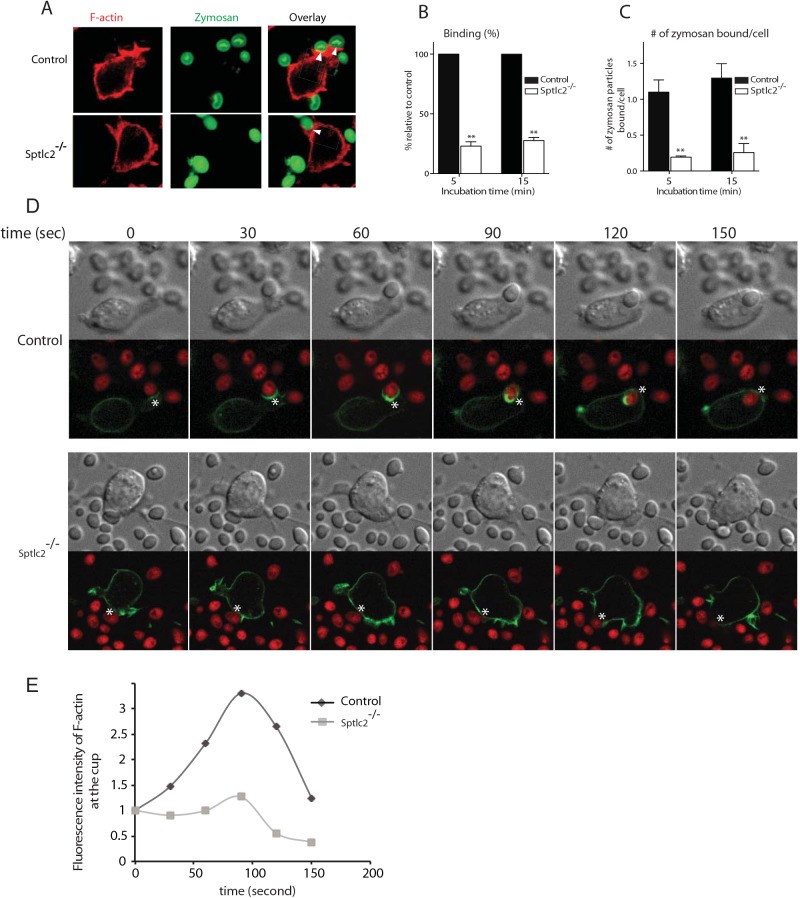
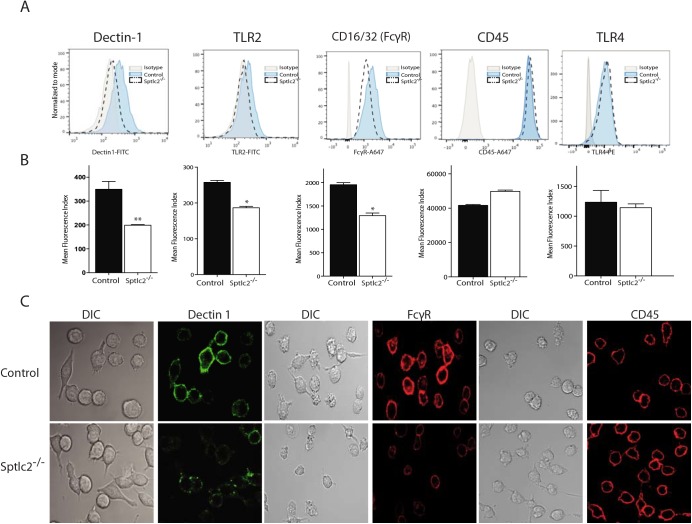
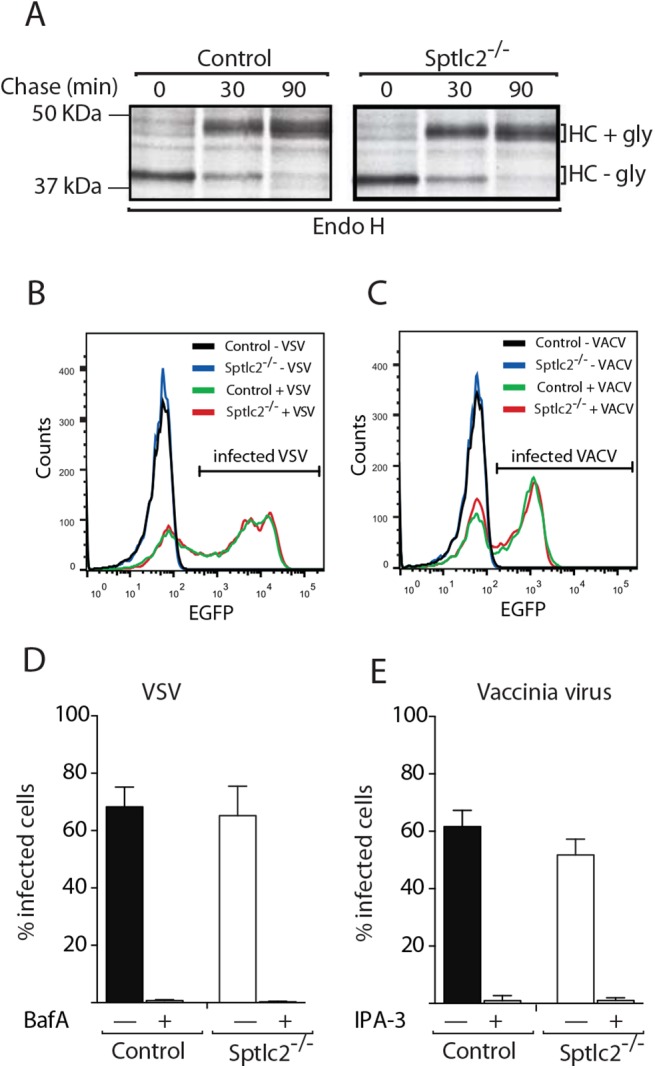
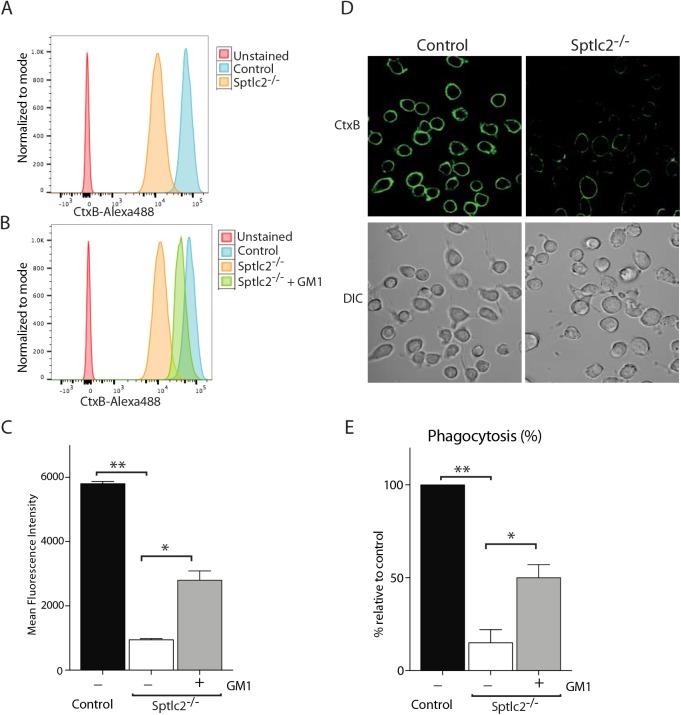
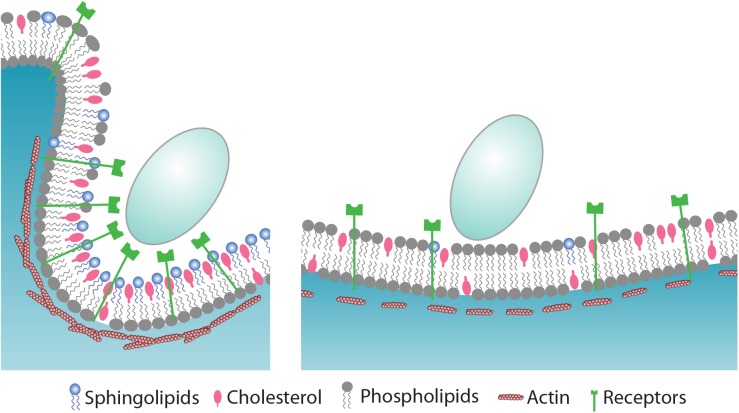
The ability of phagocytes to clear pathogens is an essential attribute of the innate immune response. The role of signaling lipid molecules such as phosphoinositides is well established, but the role of membrane sphingolipids in phagocytosis is largely unknown. Using a genetic approach and small molecule inhibitors, we show that phagocytosis of Candida albicans requires an intact sphingolipid biosynthetic pathway. Blockade of serine-palmitoyltransferase (SPT) and ceramide synthase-enzymes involved in sphingolipid biosynthesis- by myriocin and fumonisin B1, respectively, impaired phagocytosis by phagocytes. We used CRISPR/Cas9-mediated genome editing to generate Sptlc2-deficient DC2.4 dendritic cells, which lack serine palmitoyl transferase activity. Sptlc2-/- DC2.4 cells exhibited a stark defect in phagocytosis, were unable to bind fungal particles and failed to form a normal phagocytic cup to engulf C. albicans. Supplementing the growth media with GM1, the major ganglioside present at the cell surface, restored phagocytic activity of Sptlc2-/- DC2.4 cells. While overall membrane trafficking and endocytic pathways remained functional, Sptlc2-/- DC2.4 cells express reduced levels of the pattern recognition receptors Dectin-1 and TLR2 at the cell surface. Consistent with the in vitro data, compromised sphingolipid biosynthesis in mice sensitizes the animal to C. albicans infection. Sphingolipid biosynthesis is therefore critical for phagocytosis and in vivo clearance of C. albicans.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Forward genetic screen in zebrafish identifies new fungal regulators that limit host-protective Candida-innate immune interaction.mBio. 2025 May 14;16(5):e0052925. doi: 10.1128/mbio.00529-25. Epub 2025 Apr 2. mBio. 2025. PMID: 40172223 Free PMC article.
-
Phagocytes from Mice Lacking the Sts Phosphatases Have an Enhanced Antifungal Response to Candida albicans.mBio. 2018 Jul 17;9(4):e00782-18. doi: 10.1128/mBio.00782-18. mBio. 2018. PMID: 30018105 Free PMC article.
-
The Candida albicans quorum-sensing molecule farnesol alters sphingolipid metabolism in human monocyte-derived dendritic cells.mBio. 2024 Aug 14;15(8):e0073224. doi: 10.1128/mbio.00732-24. Epub 2024 Jul 2. mBio. 2024. PMID: 38953353 Free PMC article.
-
Candida albicans phospholipomannan: a sweet spot for controlling host response/inflammation.Semin Immunopathol. 2015 Mar;37(2):123-30. doi: 10.1007/s00281-014-0461-5. Epub 2014 Nov 14. Semin Immunopathol. 2015. PMID: 25394861 Review.
-
The role of pattern recognition receptors in the innate recognition of Candida albicans.Virulence. 2015;6(4):347-61. doi: 10.1080/21505594.2015.1014270. Virulence. 2015. PMID: 25714264 Free PMC article. Review.
Cited by
-
Sphingomyelin Biosynthesis Is Essential for Phagocytic Signaling during Mycobacterium tuberculosis Host Cell Entry.mBio. 2021 Jan 26;12(1):e03141-20. doi: 10.1128/mBio.03141-20. mBio. 2021. PMID: 33500344 Free PMC article.
-
Visualization and Quantification of Phagocytosis by Neutrophils.Methods Mol Biol. 2020;2087:141-148. doi: 10.1007/978-1-0716-0154-9_11. Methods Mol Biol. 2020. PMID: 31728989 Free PMC article.
-
Zymosan-Induced Murine Peritonitis Is Associated with an Increased Sphingolipid Synthesis without Changing the Long to Very Long Chain Ceramide Ratio.Int J Mol Sci. 2023 Feb 1;24(3):2773. doi: 10.3390/ijms24032773. Int J Mol Sci. 2023. PMID: 36769096 Free PMC article.
-
CRISPR-Based Genetic Manipulation of Candida Species: Historical Perspectives and Current Approaches.Front Genome Ed. 2021 Jan 8;2:606281. doi: 10.3389/fgeed.2020.606281. eCollection 2020. Front Genome Ed. 2021. PMID: 34713231 Free PMC article. Review.
-
Inhibition of sphingolipid synthesis improves outcomes and survival in GARP mutant wobbler mice, a model of motor neuron degeneration.Proc Natl Acad Sci U S A. 2020 May 12;117(19):10565-10574. doi: 10.1073/pnas.1913956117. Epub 2020 Apr 28. Proc Natl Acad Sci U S A. 2020. PMID: 32345721 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
