

Regulation of Cell Death by IAPs and Their Antagonists
- PMID: 26431568
- PMCID: PMC4861076
- DOI: 10.1016/bs.ctdb.2015.07.026
Regulation of Cell Death by IAPs and Their Antagonists
Abstract
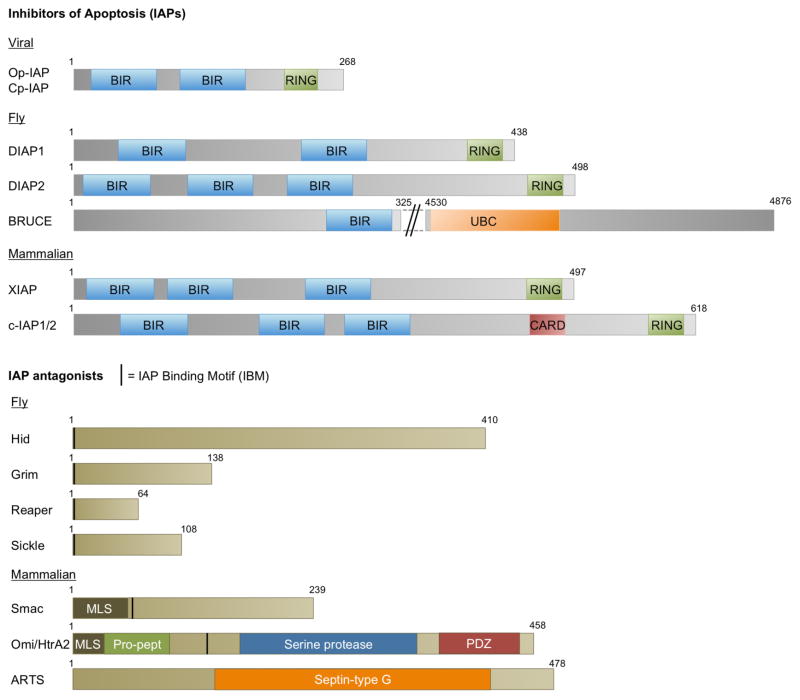
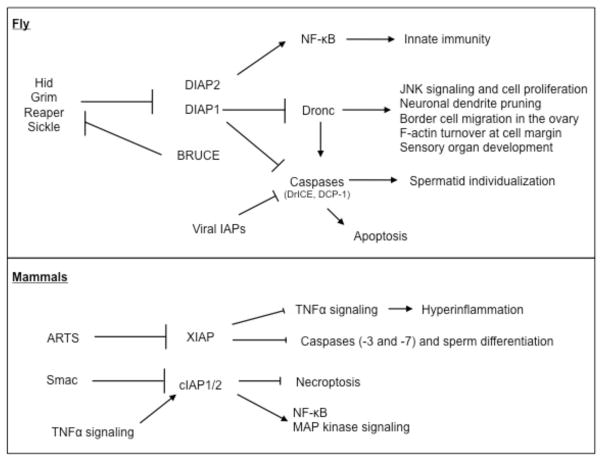
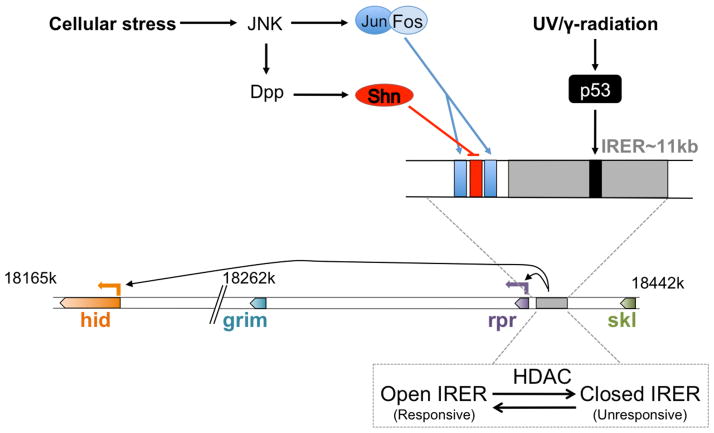
Inhibitors of apoptosis (IAPs) family of genes encode baculovirus IAP-repeat domain-containing proteins with antiapoptotic function. These proteins also contain RING or UBC domains and act by binding to major proapoptotic factors and ubiquitylating them. High levels of IAPs inhibit caspase-mediated apoptosis. For these cells to undergo apoptosis, IAP function must be neutralized by IAP-antagonists. Mammalian IAP knockouts do not exhibit obvious developmental phenotypes, but the cells are more sensitized to apoptosis in response to injury. Loss of the mammalian IAP-antagonist ARTS results in reduced stem cell apoptosis. In addition to the antiapoptotic properties, IAPs regulate the innate immune response, and the loss of IAP function in humans is associated with immunodeficiency. The roles of IAPs in Drosophila apoptosis regulation are more apparent, where the loss of IAP1, or the expression of IAP-antagonists in Drosophila cells, is sufficient to trigger apoptosis. In this organism, apoptosis as a fate is conferred by the transcriptional induction of the IAP-antagonists. Many signaling pathways often converge on shared enhancer regions of IAP-antagonists. Cell death sensitivity is further regulated by posttranscriptional mechanisms, including those regulated by kinases, miRs, and ubiquitin ligases. These mechanisms are employed to eliminate damaged or virus-infected cells, limit neuroblast (neural stem cell) numbers, generate neuronal diversity, and sculpt tissue morphogenesis.
Keywords: ARTS; Apoptosis; BRUCE; DIAP1; HID; IAP-antagonist; REAPER; SMAC; XIAP; c-IAP1.
© 2015 Elsevier Inc. All rights reserved.
Figures
References
-
- Abdelwahid E, Yokokura T, Krieser RJ, Balasundaram S, Fowle WH, White K. Mitochondrial disruption in Drosophila apoptosis. Dev Cell. 2007;12:793–806. - PubMed
-
- Akdemir F, Farkas R, Chen P, Juhasz G, Medved’ova L, Sass M, Wang L, Wang X, Chittaranjan S, Gorski SM, Rodriguez A, Abrams JM. Autophagy occurs upstream or parallel to the apoptosome during histolytic cell death. Development. 2006;113:1457–1465. - PubMed
-
- Arama E, Agapite J, Steller H. Caspase activity and a specific cytochrome C are required for sperm differentiation in Drosophila. Dev Cell. 2003;4:687–697. - PubMed
-
- Bartke T, Pohl C, Pyrowolakis G, Jentsch S. Dual role of BRUCE as an antiapoptotic IAP and a chimeric E2/E3 ubiquitin ligase. Mol Cell. 2004;14:801–811. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
