

The EGFR-HER2 module: a stem cell approach to understanding a prime target and driver of solid tumors
- PMID: 26434585
- PMCID: PMC4820040
- DOI: 10.1038/onc.2015.372
The EGFR-HER2 module: a stem cell approach to understanding a prime target and driver of solid tumors
Abstract
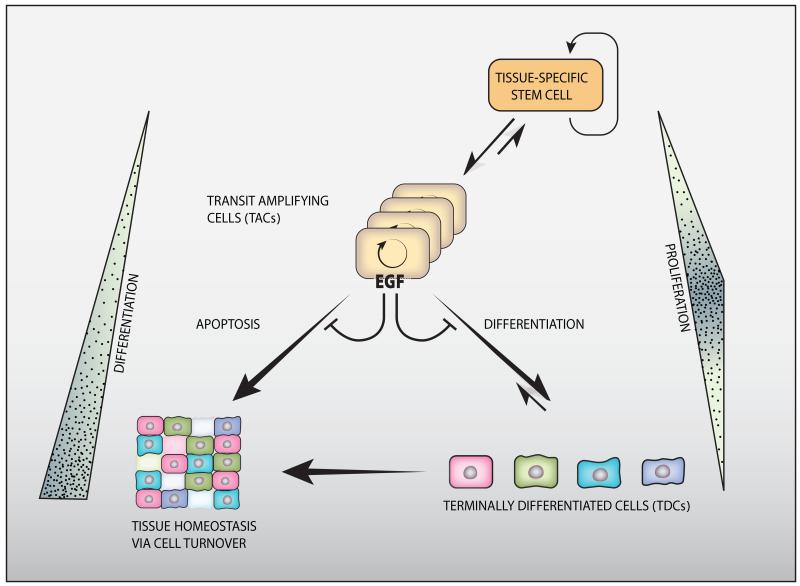
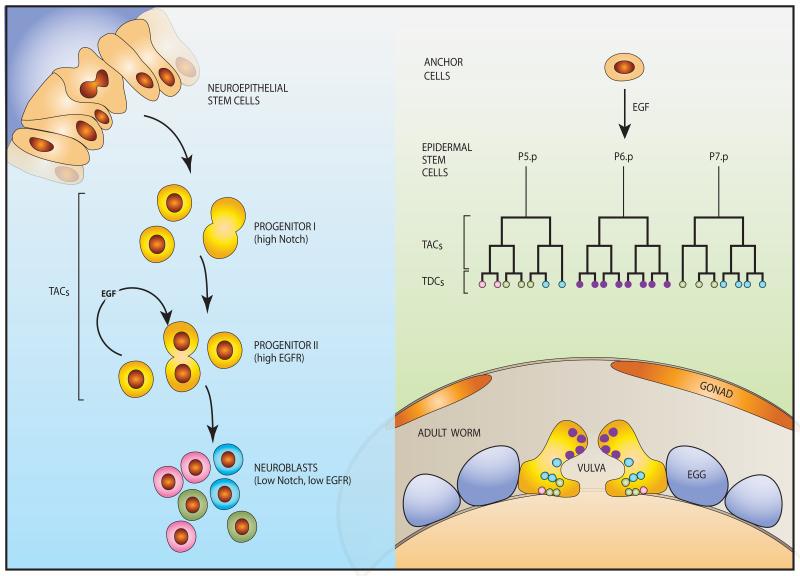
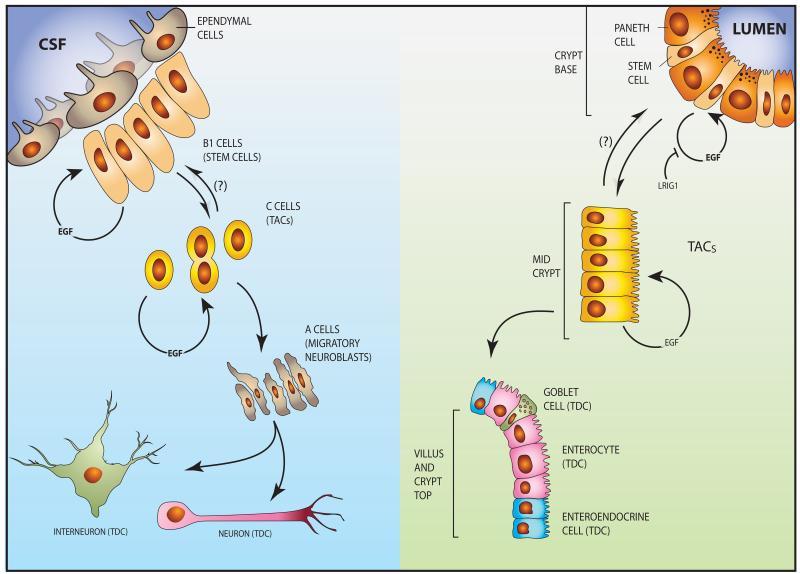
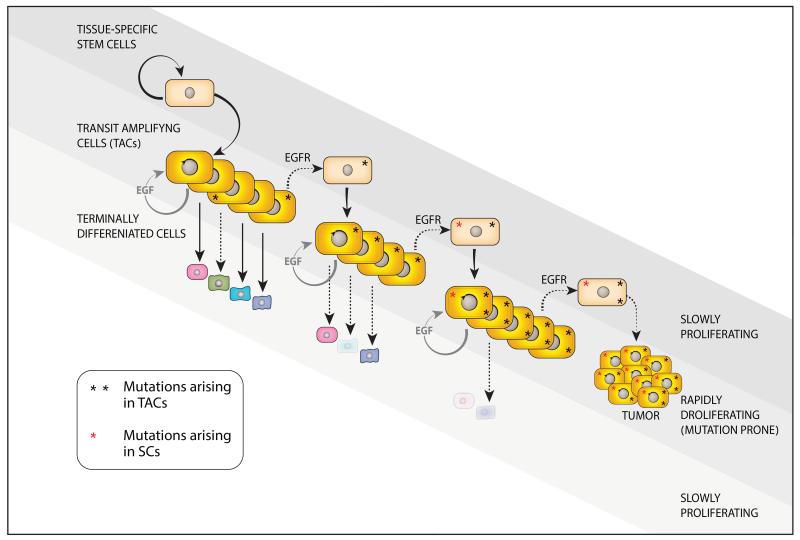
The epidermal growth factor receptor (EGFR) and a coreceptor denoted HER2/ERBB2 are frequently overexpressed or mutated in solid tumors, such as carcinomas and gliomas. In line with driver roles, cancer drugs intercepting EGFR or HER2 currently outnumber therapies targeting other hubs of signal transduction. To explain the roles for EGFR and HER2 as prime drivers and targets, we take lessons from invertebrates and refer to homeostatic regulation of several mammalian tissues. The model we infer ascribes to the EGFR-HER2 module pivotal functions in rapid clonal expansion of progenitors called transient amplifying cells (TACs). Accordingly, TACs of tumors suffer from replication stress, and hence accumulate mutations. In addition, several lines of evidence propose that in response to EGF and related mitogens, TACs might undergo dedifferentiation into tissue stem cells, which might enable entry of oncogenic mutations into the stem cell compartment. According to this view, antibodies or kinase inhibitors targeting EGFR-HER2 effectively retard some solid tumors because they arrest mutation-enriched TACs and possibly inhibit their dedifferentiation. Deeper understanding of the EGFR-HER2 module and relations between cancer stem cells and TACs will enhance our ability to control a broad spectrum of human malignancies.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
