

Mechanism and regulation of cytoplasmic dynein
- PMID: 26436706
- PMCID: PMC4644480
- DOI: 10.1146/annurev-cellbio-100814-125438
Mechanism and regulation of cytoplasmic dynein
Abstract
Until recently, dynein was the least understood of the cytoskeletal motors. However, a wealth of new structural, mechanistic, and cell biological data is shedding light on how this complicated minus-end-directed, microtubule-based motor works. Cytoplasmic dynein-1 performs a wide array of functions in most eukaryotes, both in interphase, in which it transports organelles, proteins, mRNAs, and viruses, and in mitosis and meiosis. Mutations in dynein or its regulators are linked to neurodevelopmental and neurodegenerative diseases. Here, we begin by providing a synthesis of recent data to describe the current model of dynein's mechanochemical cycle. Next, we discuss regulators of dynein, with particular focus on those that directly interact with the motor to modulate its recruitment to microtubules, initiate cargo transport, or activate minus-end-directed motility.
Keywords: Lis1; Nudel/NudE; cytoskeleton; dynactin; microtubule; transport.
Figures
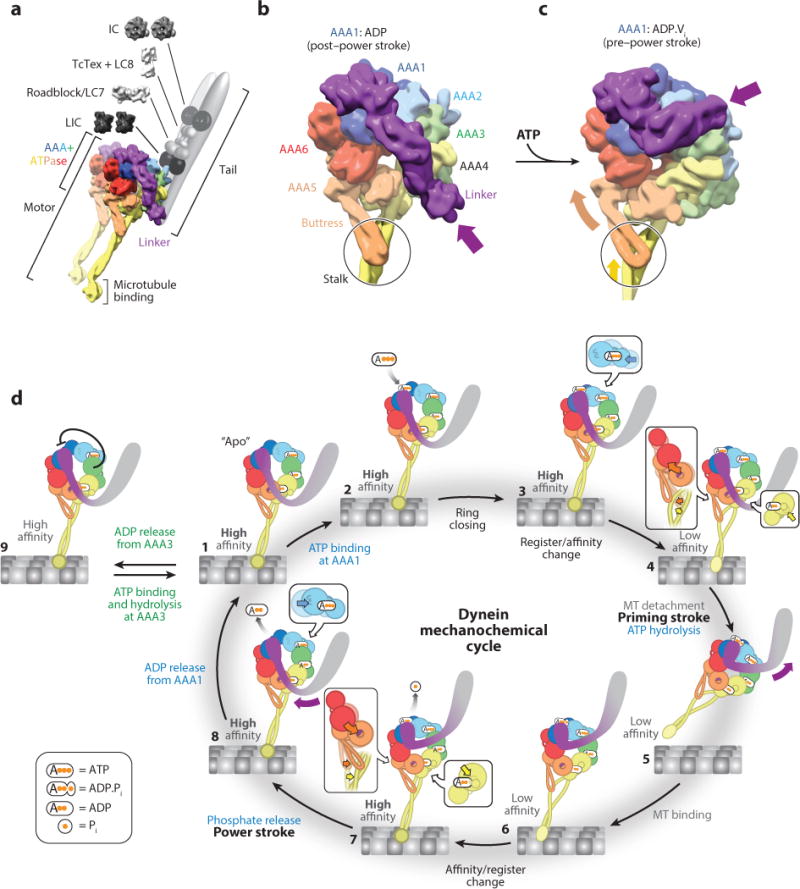
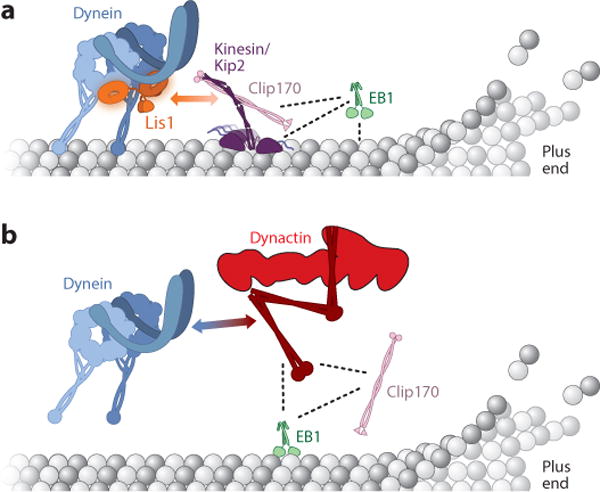
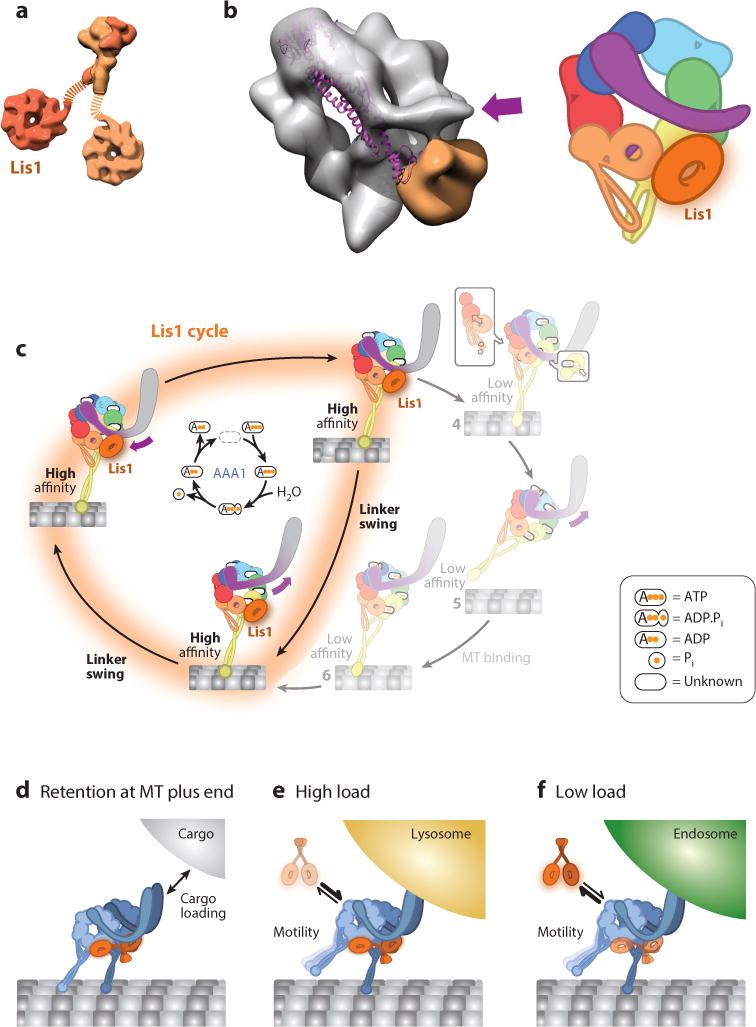
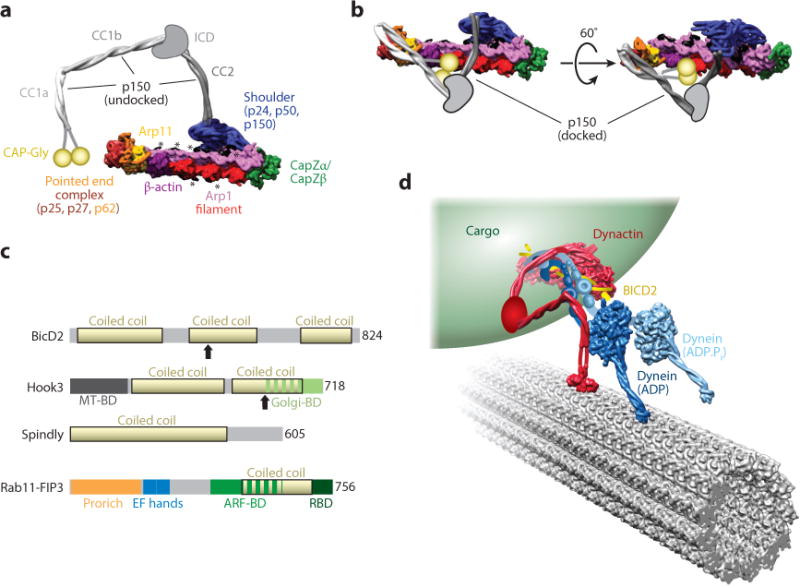
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
