

cKit+ cardiac progenitors of neural crest origin
- PMID: 26438843
- PMCID: PMC4620867
- DOI: 10.1073/pnas.1517201112
cKit+ cardiac progenitors of neural crest origin
Abstract
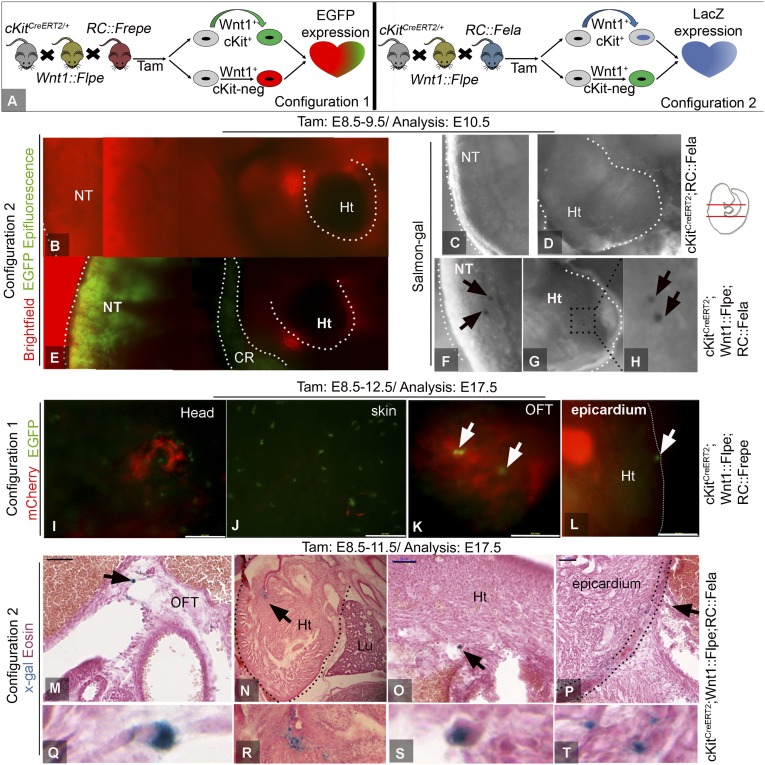
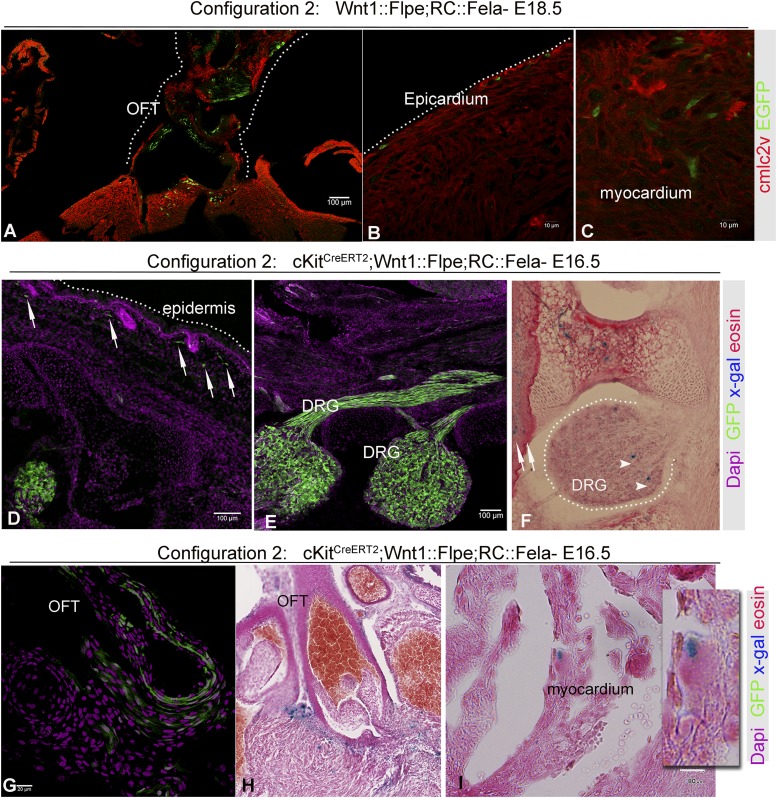
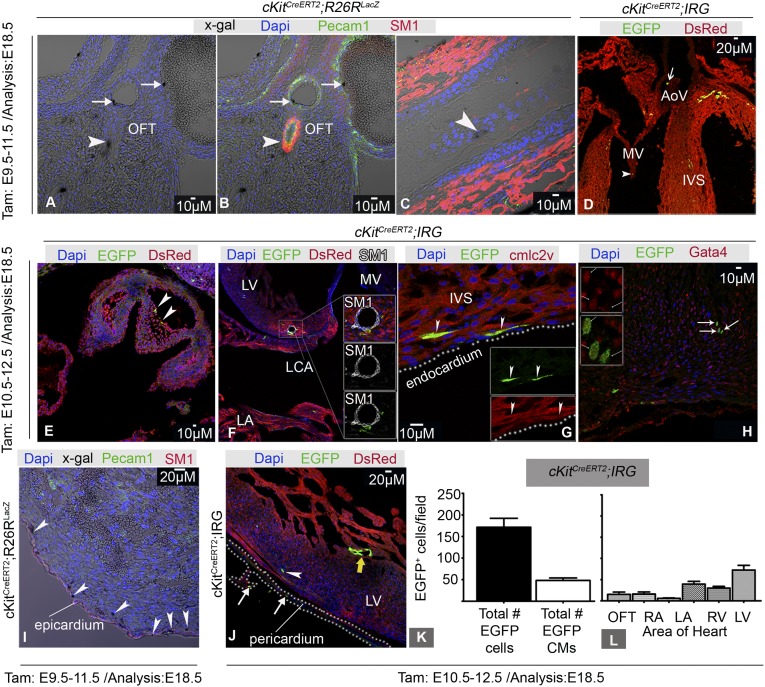
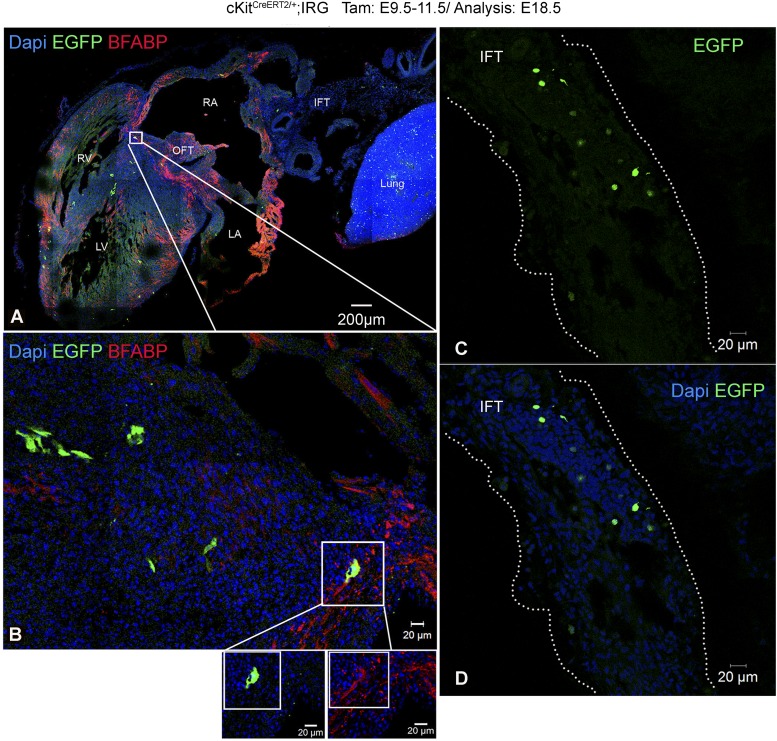
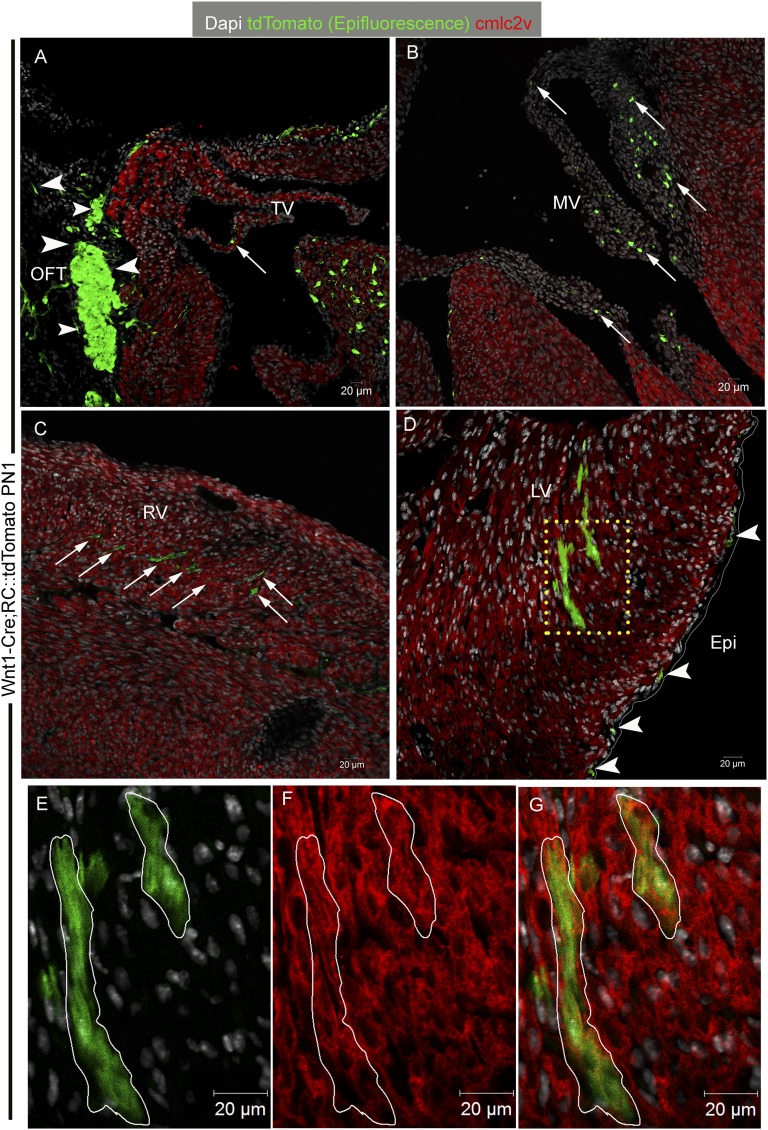
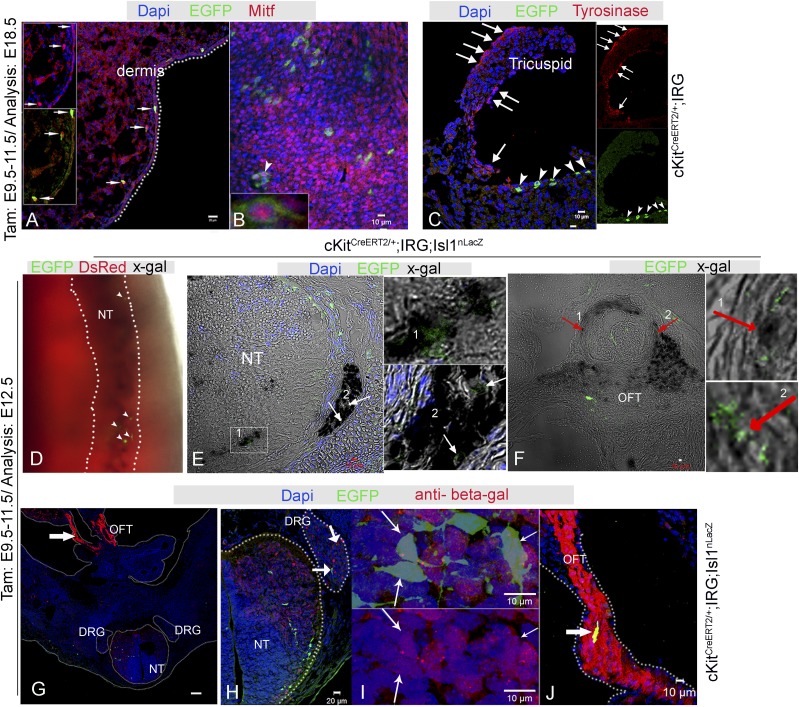
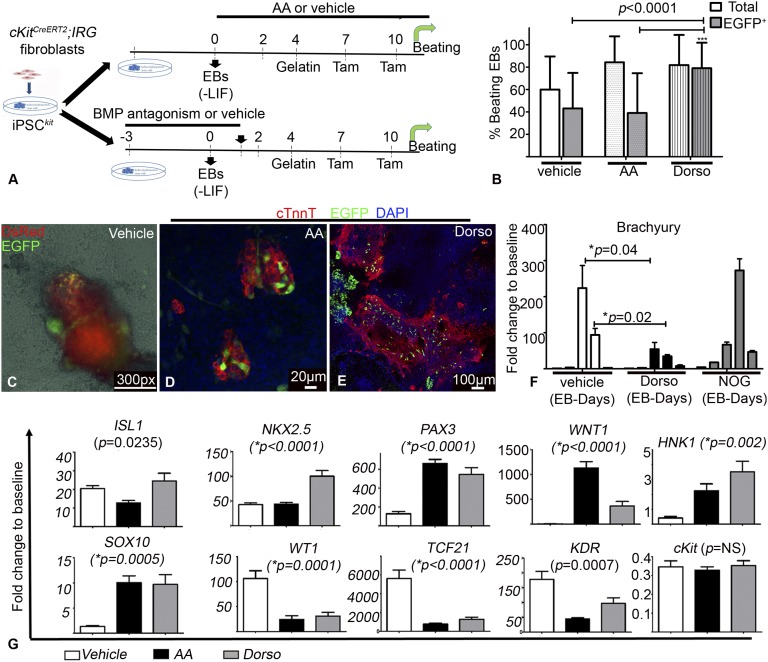
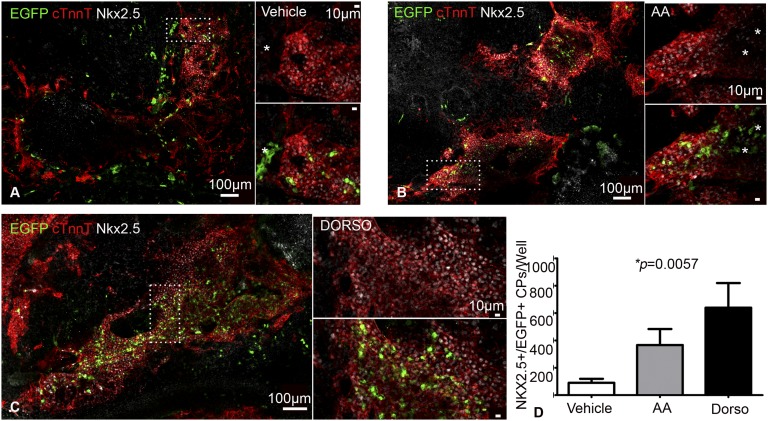
The degree to which cKit-expressing progenitors generate cardiomyocytes in the heart is controversial. Genetic fate-mapping studies suggest minimal contribution; however, whether or not minimal contribution reflects minimal cardiomyogenic capacity is unclear because the embryonic origin and role in cardiogenesis of these progenitors remain elusive. Using high-resolution genetic fate-mapping approaches with cKit(CreERT2/+) and Wnt1::Flpe mouse lines, we show that cKit delineates cardiac neural crest progenitors (CNC(kit)). CNC(kit) possess full cardiomyogenic capacity and contribute to all CNC derivatives, including cardiac conduction system cells. Furthermore, by modeling cardiogenesis in cKit(CreERT2)-induced pluripotent stem cells, we show that, paradoxically, the cardiogenic fate of CNC(kit) is regulated by bone morphogenetic protein antagonism, a signaling pathway activated transiently during establishment of the cardiac crescent, and extinguished from the heart before CNC invasion. Together, these findings elucidate the origin of cKit(+) cardiac progenitors and suggest that a nonpermissive cardiac milieu, rather than minimal cardiomyogenic capacity, controls the degree of CNC(kit) contribution to myocardium.
Keywords: BMP signalling; cardiac neural crest; cardiac stem cells; cardiomyogenesis.
Conflict of interest statement
Conflict of interest statement: K.E.H. and J.M.H. report having a patent for cardiac cell-based therapy. K.E.H. and J.M.H. own equity in Vestion Inc. and are members of the scientific advisory board and consultants of Vestion, Inc. J.M.H. is a board member of Vestion Inc. Vestion Inc. did not participate in funding this work. The other authors report no conflicts.
Figures
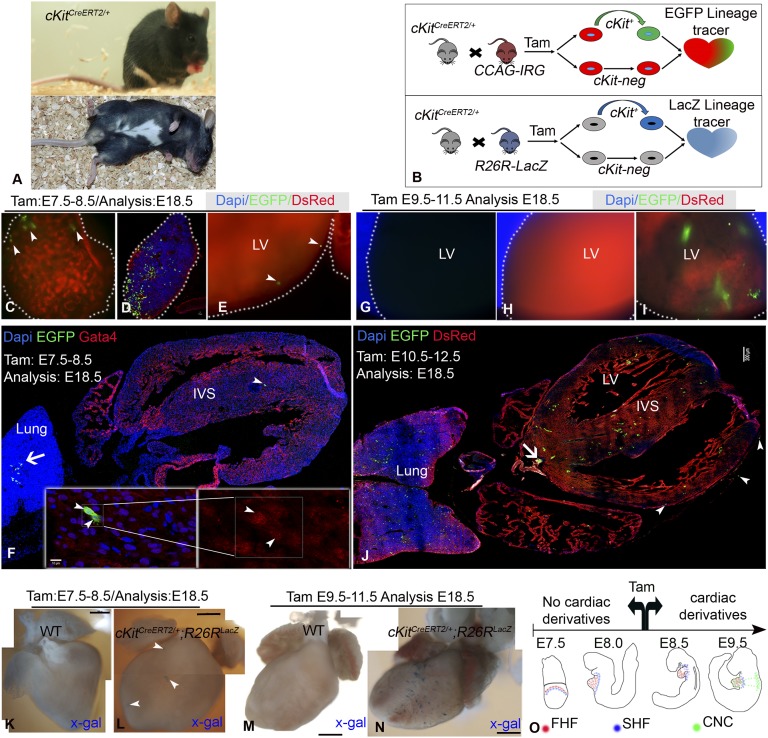
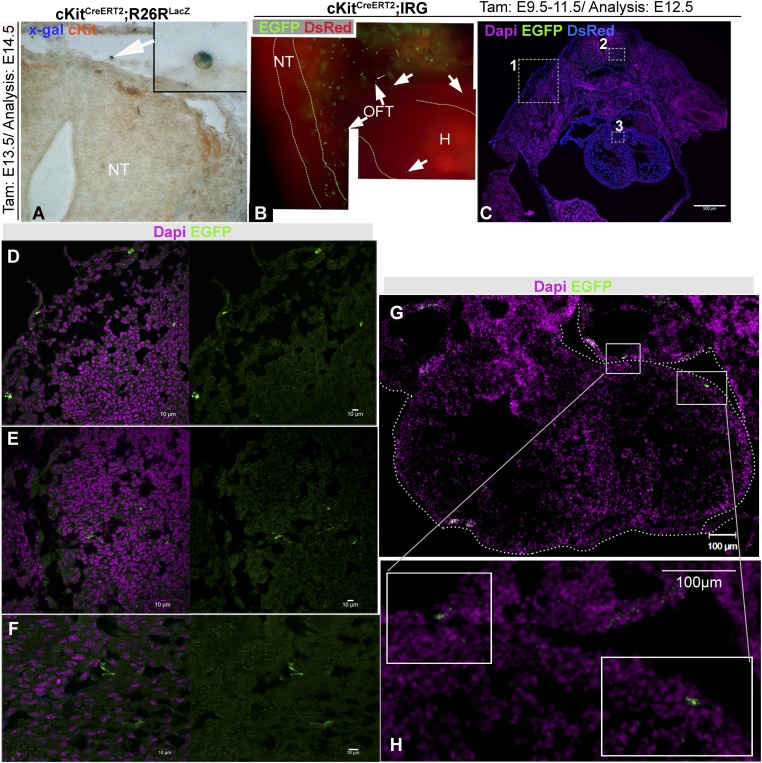
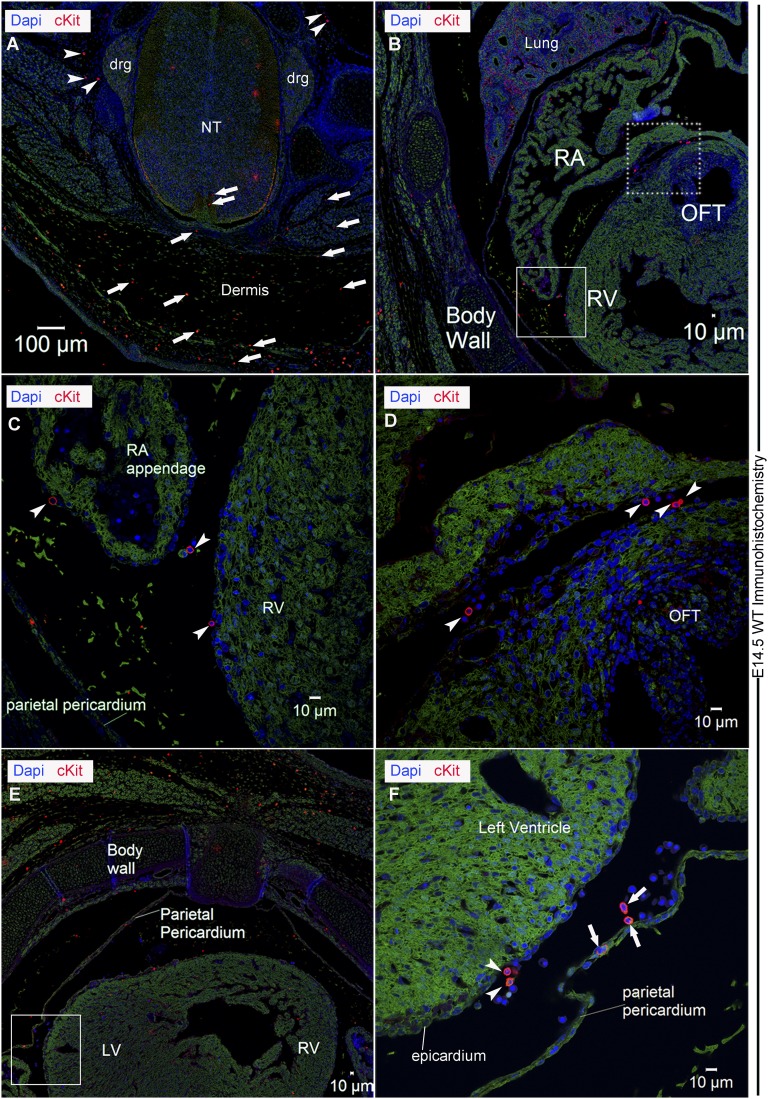
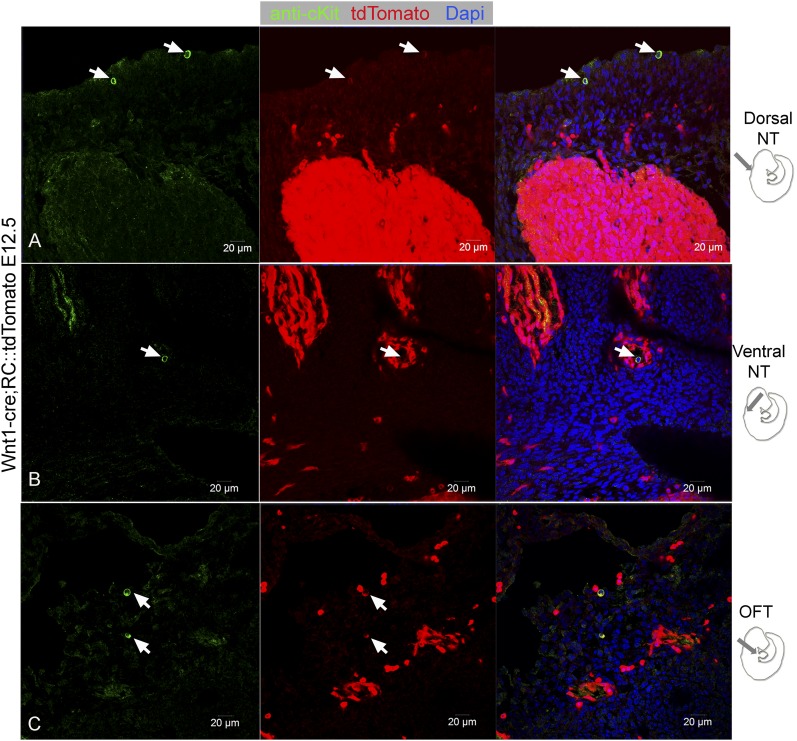
Comment in
-
c-kit+ Cardiac Stem Cells: Spontaneous Creation or a Perplexing Reality.Circ Res. 2016 Mar 4;118(5):783-5. doi: 10.1161/CIRCRESAHA.115.308103. Circ Res. 2016. PMID: 26941422 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
