

The Genomic Landscape and Clinical Relevance of A-to-I RNA Editing in Human Cancers
- PMID: 26439496
- PMCID: PMC4605878
- DOI: 10.1016/j.ccell.2015.08.013
The Genomic Landscape and Clinical Relevance of A-to-I RNA Editing in Human Cancers
Abstract
Adenosine-to-inosine (A-to-I) RNA editing is a widespread post-transcriptional mechanism, but its genomic landscape and clinical relevance in cancer have not been investigated systematically. We characterized the global A-to-I RNA editing profiles of 6,236 patient samples of 17 cancer types from The Cancer Genome Atlas and revealed a striking diversity of altered RNA-editing patterns in tumors relative to normal tissues. We identified an appreciable number of clinically relevant editing events, many of which are in noncoding regions. We experimentally demonstrated the effects of several cross-tumor nonsynonymous RNA editing events on cell viability and provide the evidence that RNA editing could selectively affect drug sensitivity. These results highlight RNA editing as an exciting theme for investigating cancer mechanisms, biomarkers, and treatments.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
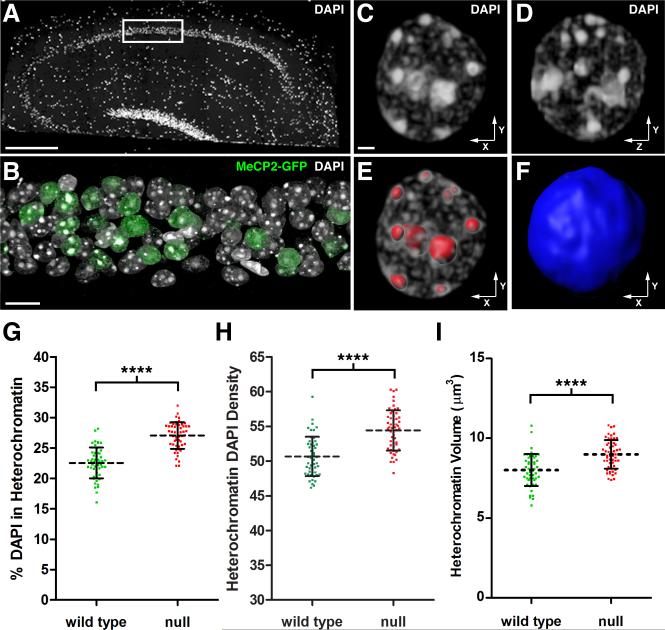
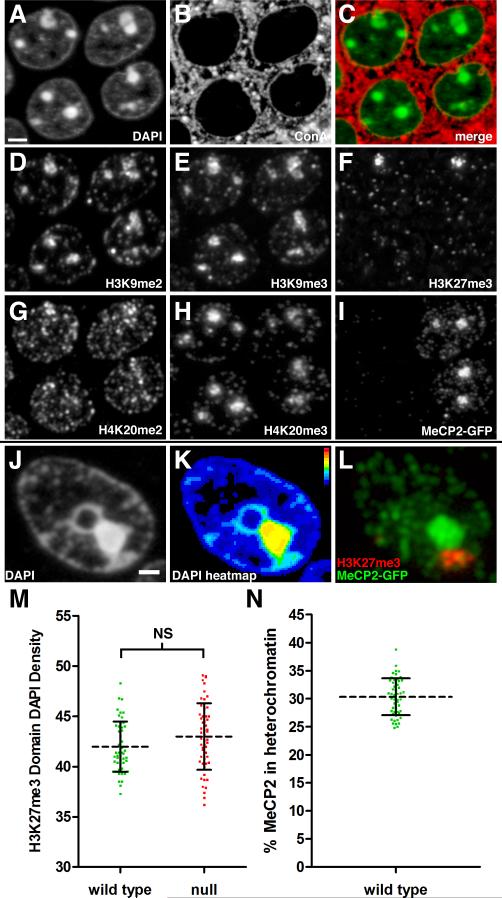
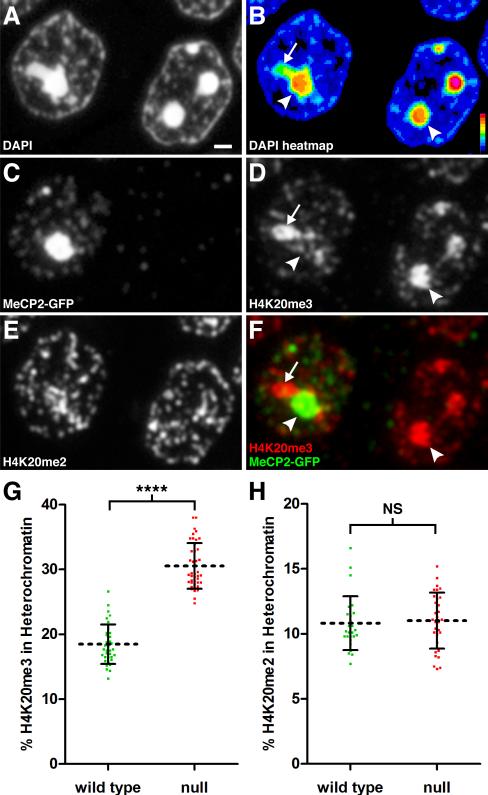
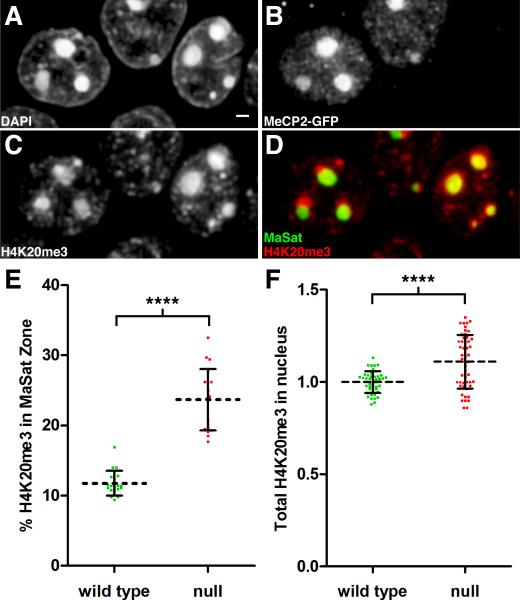
Comment in
-
Cancer genomics: The relevance of extensive editing in tumour transcriptomes.Nat Rev Cancer. 2015 Dec;15(12):698. doi: 10.1038/nrc4044. Epub 2015 Oct 29. Nat Rev Cancer. 2015. PMID: 26511457 No abstract available.
-
RNA Editing Dynamically Rewrites the Cancer Code.Trends Cancer. 2015 Dec;1(4):211-212. doi: 10.1016/j.trecan.2015.10.008. Epub 2015 Nov 26. Trends Cancer. 2015. PMID: 27695712 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
- P30 CA016672/CA/NCI NIH HHS/United States
- R01GM102484/GM/NIGMS NIH HHS/United States
- CA016672/CA/NCI NIH HHS/United States
- P50 CA098258/CA/NCI NIH HHS/United States
- U01 CA168394/CA/NCI NIH HHS/United States
- U24CA143883/CA/NCI NIH HHS/United States
- CA098258/CA/NCI NIH HHS/United States
- R01CA175486/CA/NCI NIH HHS/United States
- P50 CA083639/CA/NCI NIH HHS/United States
- R01 GM102484/GM/NIGMS NIH HHS/United States
- R01 CA175486/CA/NCI NIH HHS/United States
- U24 CA143883/CA/NCI NIH HHS/United States
- CA168394/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
