

Microhomology-Mediated End Joining: A Back-up Survival Mechanism or Dedicated Pathway?
- PMID: 26439531
- PMCID: PMC4638128
- DOI: 10.1016/j.tibs.2015.08.006
Microhomology-Mediated End Joining: A Back-up Survival Mechanism or Dedicated Pathway?
Abstract
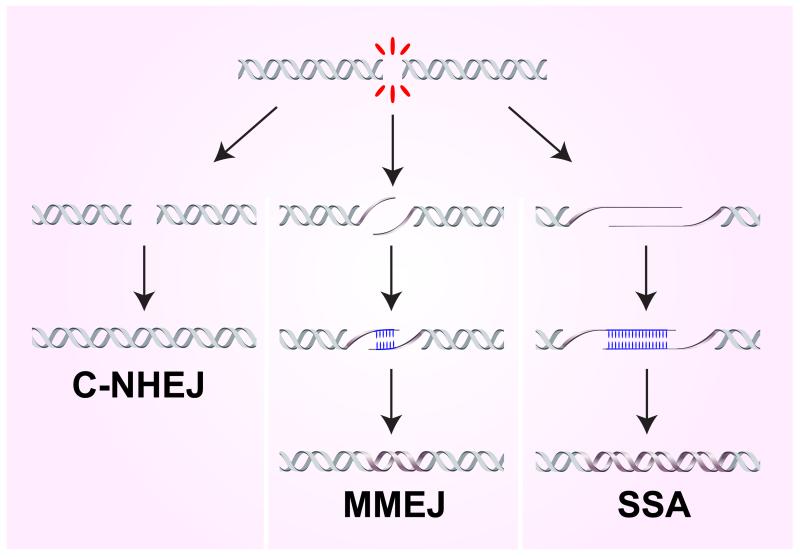
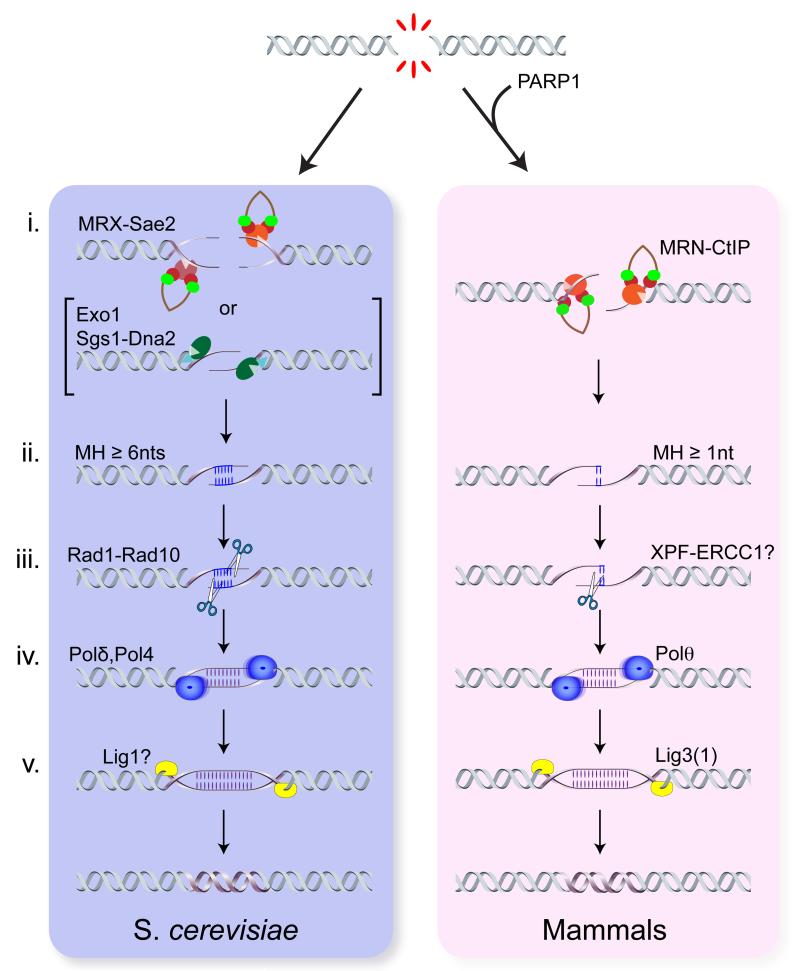
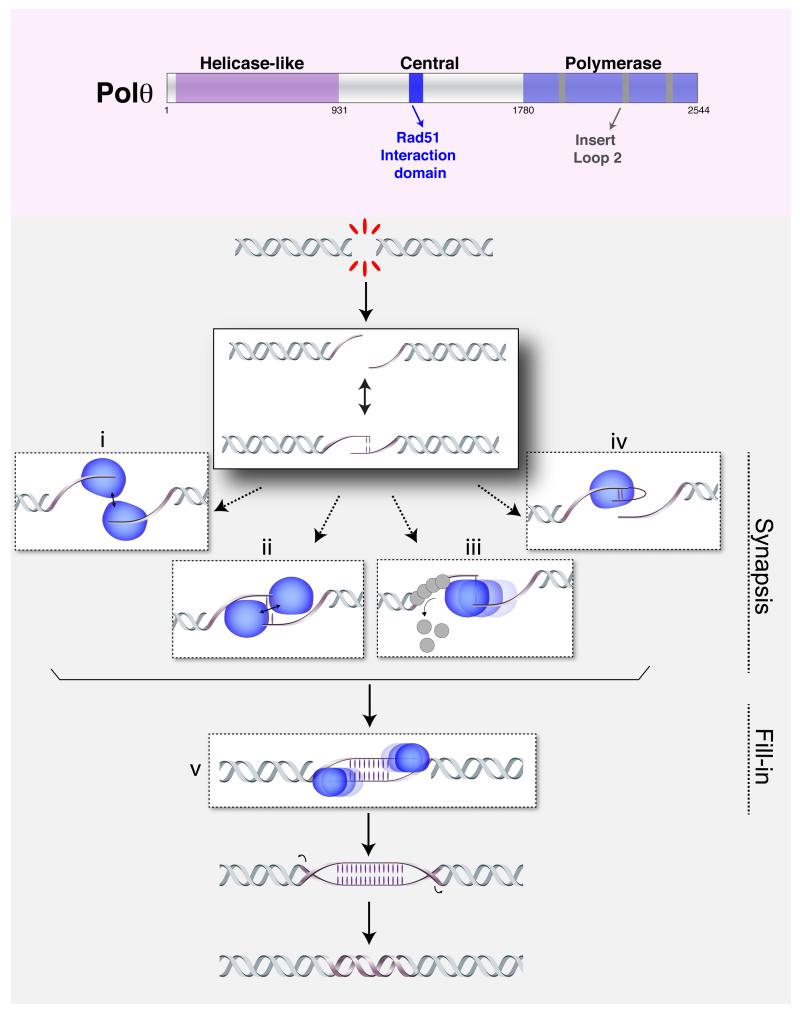
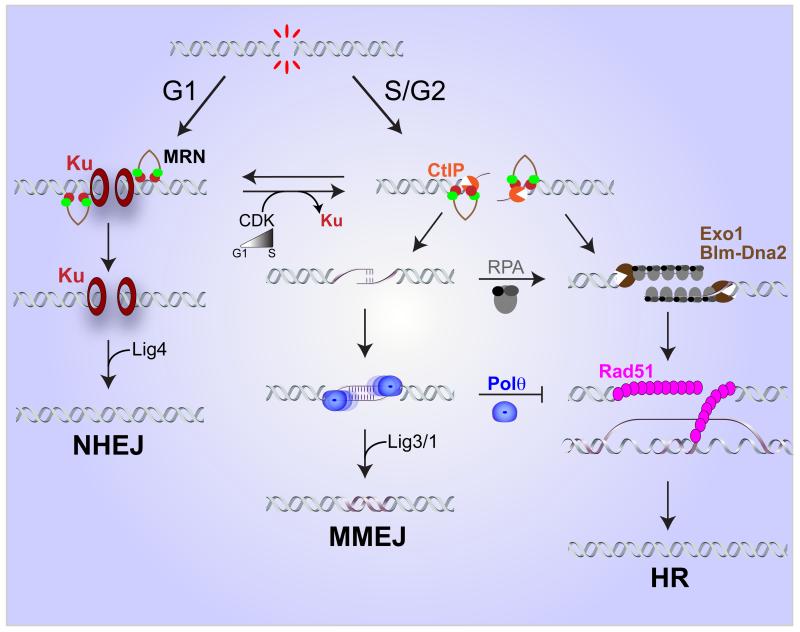
DNA double-strand breaks (DSBs) disrupt the continuity of chromosomes and their repair by error-free mechanisms is essential to preserve genome integrity. Microhomology-mediated end joining (MMEJ) is an error-prone repair mechanism that involves alignment of microhomologous sequences internal to the broken ends before joining, and is associated with deletions and insertions that mark the original break site, as well as chromosome translocations. Whether MMEJ has a physiological role or is simply a back-up repair mechanism is a matter of debate. Here we review recent findings pertaining to the mechanism of MMEJ and discuss its role in normal and cancer cells.
Keywords: DNA Polθ; MMEJ; chromosomal translocations; end joining; homologous recombination; microhomology.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
