

All spiking, sustained ON displaced amacrine cells receive gap-junction input from melanopsin ganglion cells
- PMID: 26441349
- PMCID: PMC4631663
- DOI: 10.1016/j.cub.2015.09.018
All spiking, sustained ON displaced amacrine cells receive gap-junction input from melanopsin ganglion cells
Erratum in
- Curr Biol. 2015 Nov 2;25(21):2878
Abstract
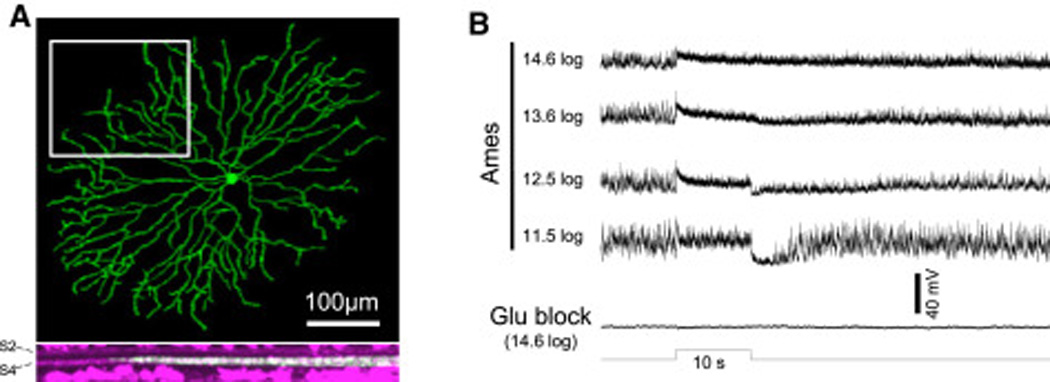
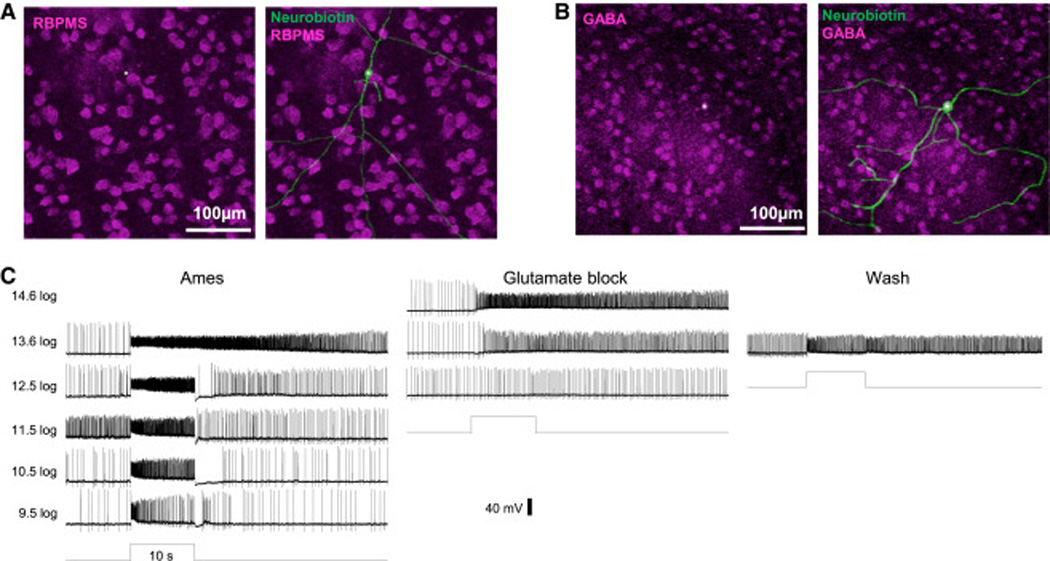
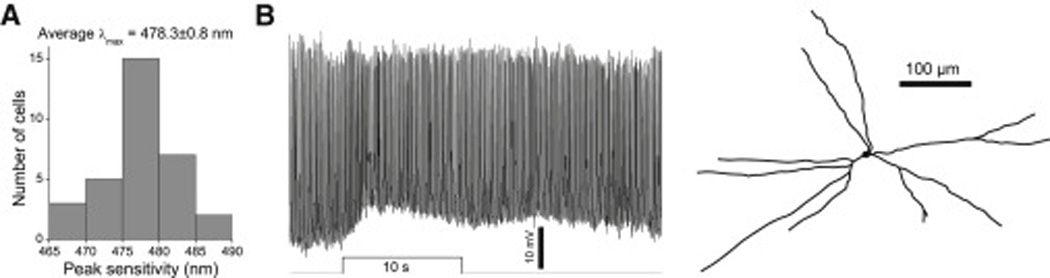
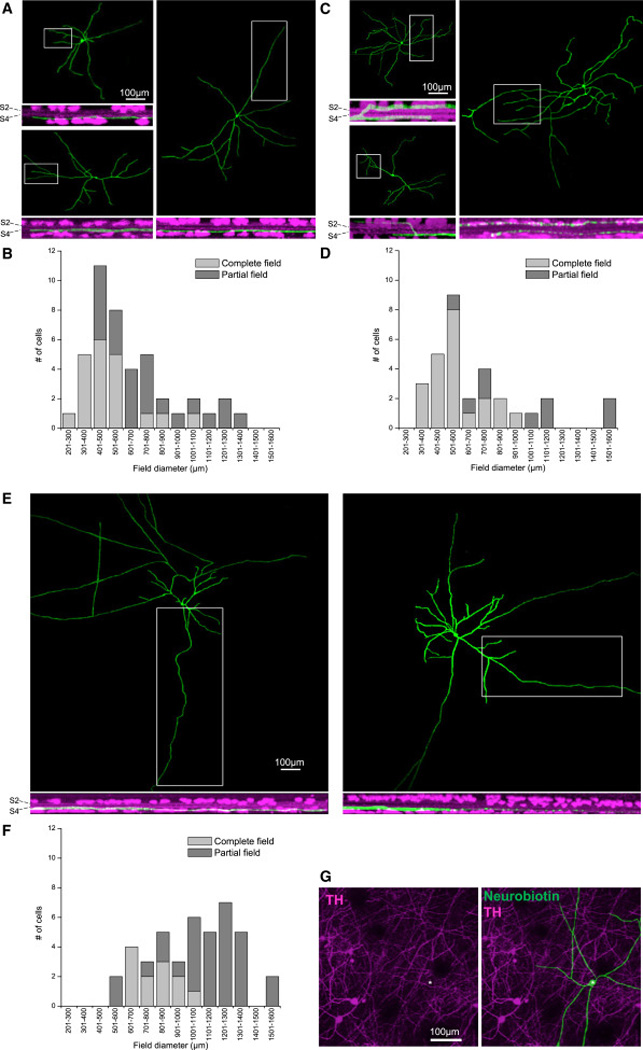
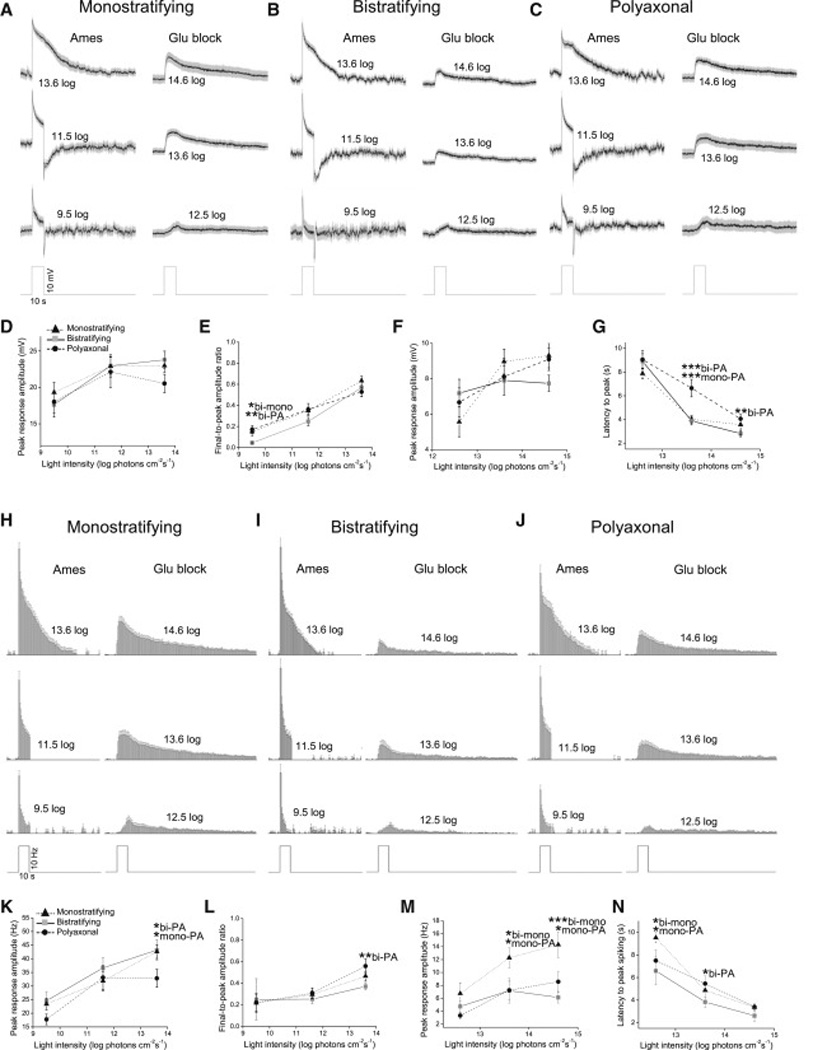
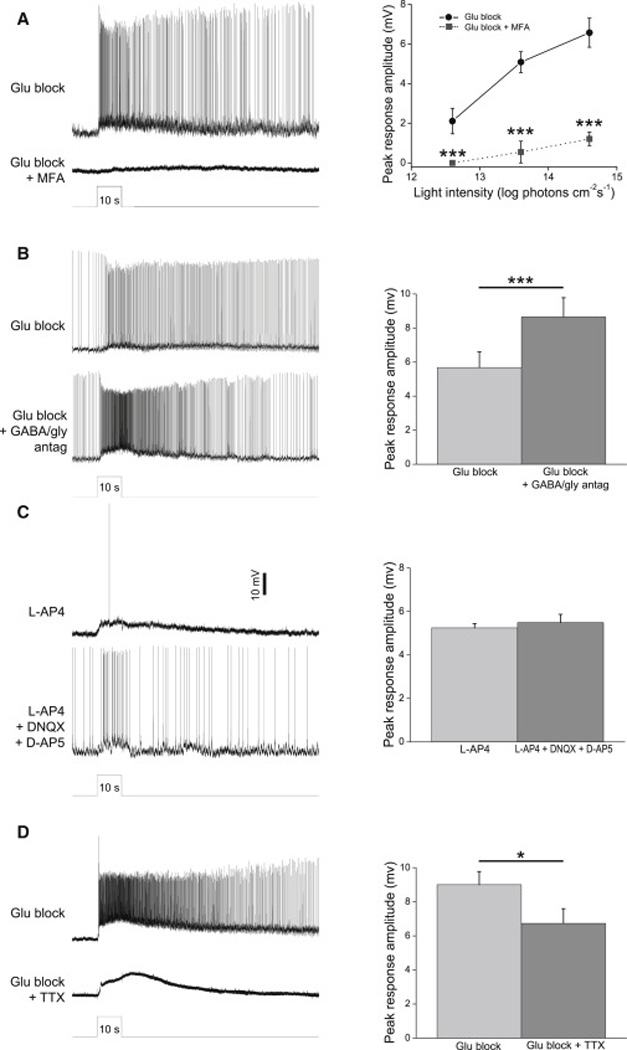
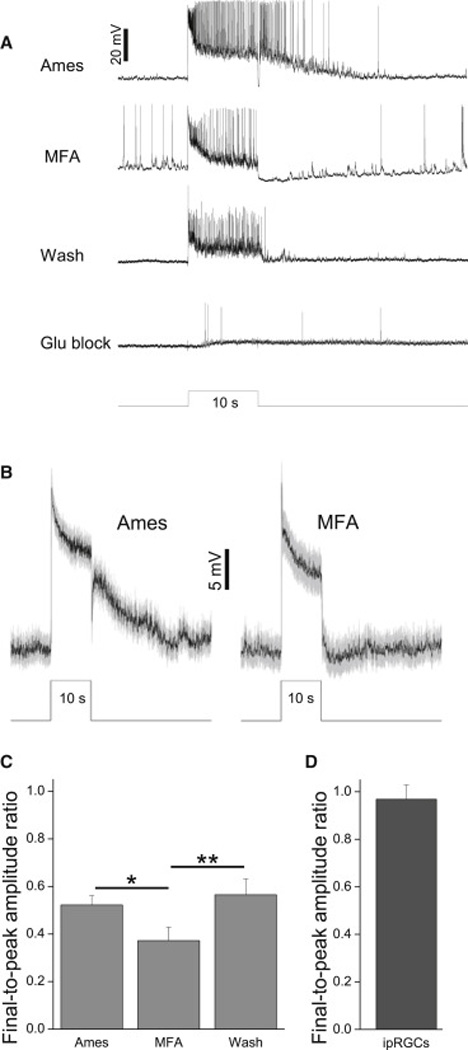
Retinal neurons exhibit sustained versus transient light responses, which are thought to encode low- and high-frequency stimuli, respectively. This dichotomy has been recognized since the earliest intracellular recordings from the 1960s, but the underlying mechanisms are not yet fully understood. We report that in the ganglion cell layer of rat retinas, all spiking amacrine interneurons with sustained ON photoresponses receive gap-junction input from intrinsically photosensitive retinal ganglion cells (ipRGCs), recently discovered photoreceptors that specialize in prolonged irradiance detection. This input presumably allows ipRGCs to regulate the secretion of neuromodulators from these interneurons. We have identified three morphological varieties of such ipRGC-driven displaced amacrine cells: (1) monostratified cells with dendrites terminating exclusively in sublamina S5 of the inner plexiform layer, (2) bistratified cells with dendrites in both S1 and S5, and (3) polyaxonal cells with dendrites and axons stratifying in S5. Most of these amacrine cells are wide field, although some are medium field. The three classes respond to light differently, suggesting that they probably perform diverse functions. These results demonstrate that ipRGCs are a major source of tonic visual information within the retina and exert widespread intraretinal influence. They also add to recent evidence that ganglion cells signal not only to the brain.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures
References
-
- Masland RH. The fundamental plan of the retina. Nat Neurosci. 2001;4:877–886. - PubMed
-
- Dong CJ, Werblin FS. Temporal contrast enhancement via GABAC feedback at bipolar terminals in the tiger salamander retina. J Neurophysiol. 1998;79:2171–2180. - PubMed
-
- Maguire G. Rapid desensitization converts prolonged glutamate release into a transient EPSC at ribbon synapses between retinal bipolar and amacrine cells. Eur J Neurosci. 1999;11:353–362. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
