

Ontogeny of Tissue-Resident Macrophages
- PMID: 26441990
- PMCID: PMC4585135
- DOI: 10.3389/fimmu.2015.00486
Ontogeny of Tissue-Resident Macrophages
Abstract
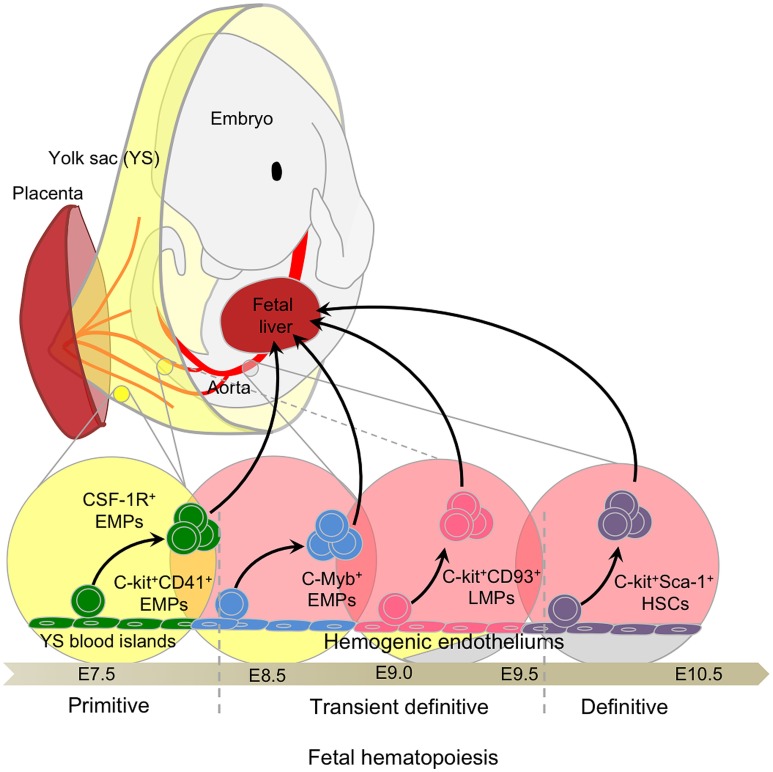
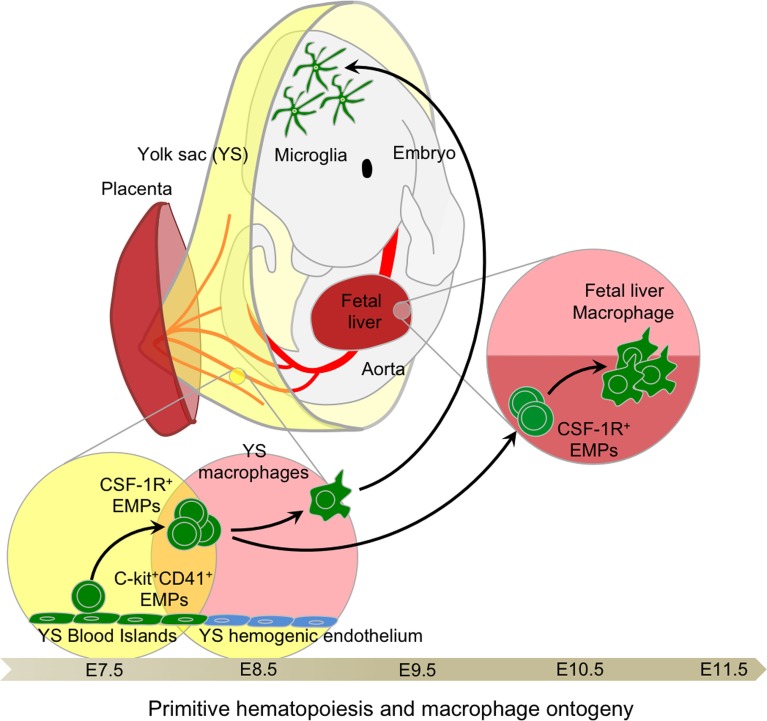
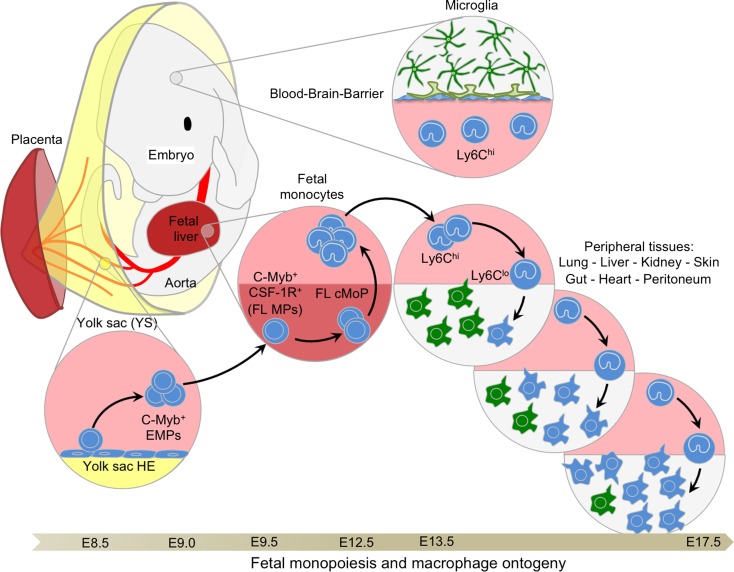
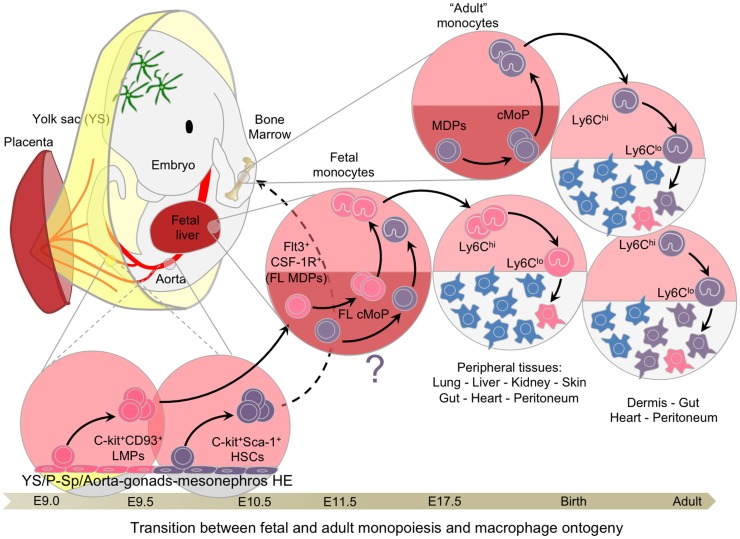
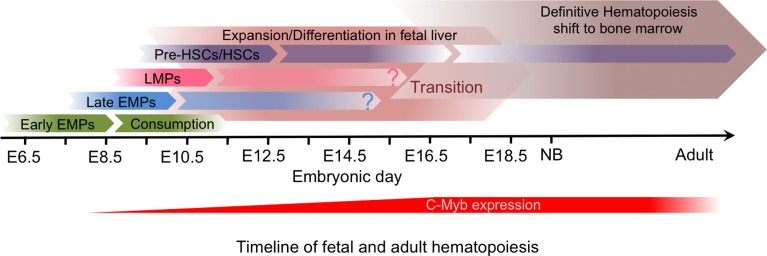
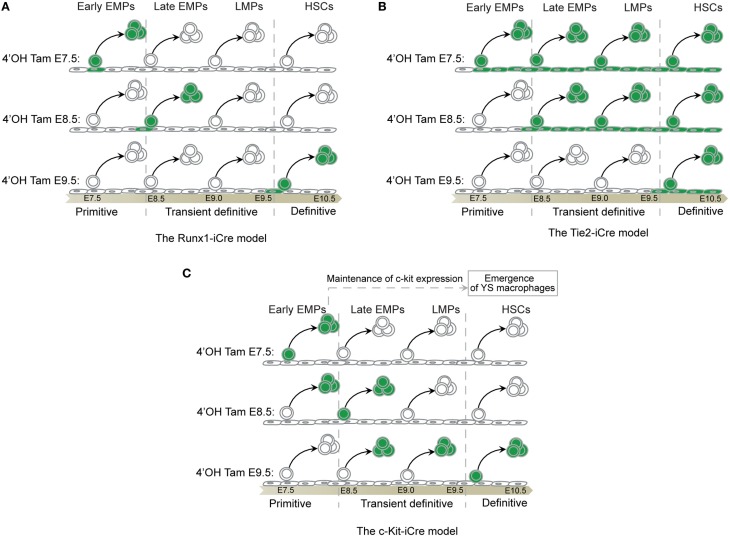
The origin of tissue-resident macrophages, crucial for homeostasis and immunity, has remained controversial until recently. Originally described as part of the mononuclear phagocyte system, macrophages were long thought to derive solely from adult blood circulating monocytes. However, accumulating evidence now shows that certain macrophage populations are in fact independent from monocyte and even from adult bone marrow hematopoiesis. These tissue-resident macrophages derive from sequential seeding of tissues by two precursors during embryonic development. Primitive macrophages generated in the yolk sac (YS) from early erythro-myeloid progenitors (EMPs), independently of the transcription factor c-Myb and bypassing monocytic intermediates, first give rise to microglia. Later, fetal monocytes, generated from c-Myb(+) EMPs that initially seed the fetal liver (FL), then give rise to the majority of other adult macrophages. Thus, hematopoietic stem cell-independent embryonic precursors transiently present in the YS and the FL give rise to long-lasting self-renewing macrophage populations.
Keywords: C-Myb; erythro-myeloid progenitors; fetal liver; hematopoiesis; hematopoietic stem cells; macrophages; monocytes; yolk sac.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources