

The different facets of organelle interplay-an overview of organelle interactions
- PMID: 26442263
- PMCID: PMC4585249
- DOI: 10.3389/fcell.2015.00056
The different facets of organelle interplay-an overview of organelle interactions
Abstract
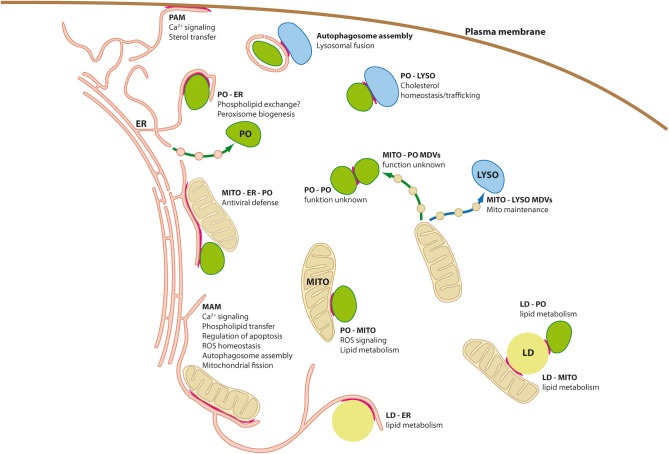
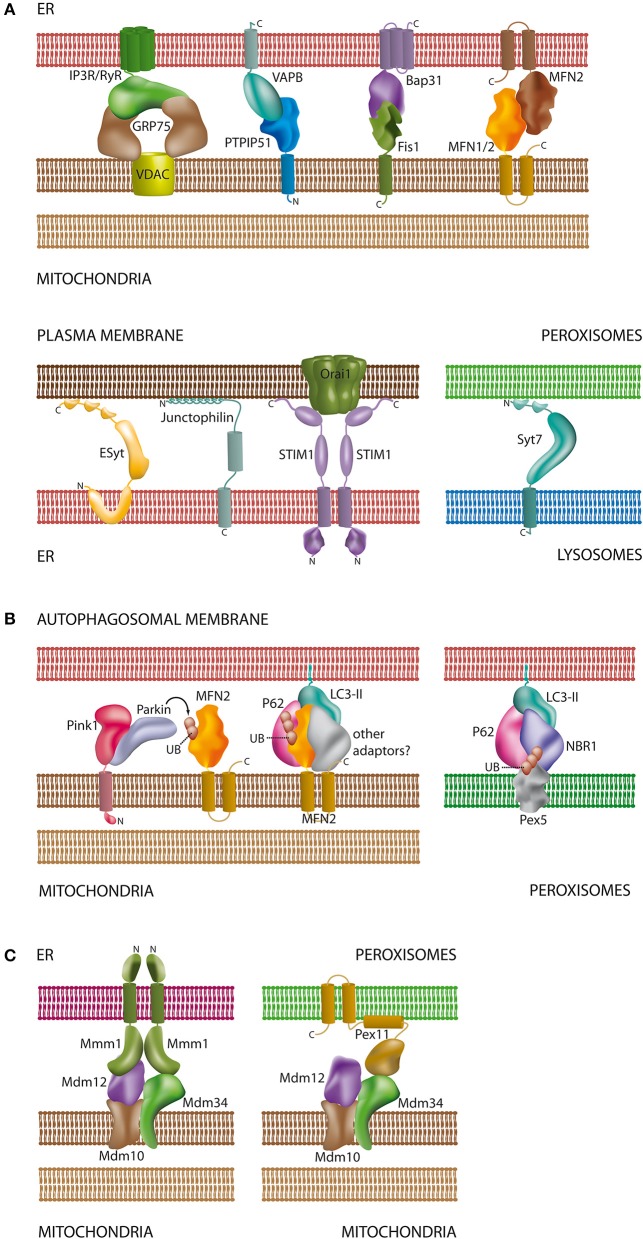
Membrane-bound organelles such as mitochondria, peroxisomes, or the endoplasmic reticulum (ER) create distinct environments to promote specific cellular tasks such as ATP production, lipid breakdown, or protein export. During recent years, it has become evident that organelles are integrated into cellular networks regulating metabolism, intracellular signaling, cellular maintenance, cell fate decision, and pathogen defence. In order to facilitate such signaling events, specialized membrane regions between apposing organelles bear distinct sets of proteins to enable tethering and exchange of metabolites and signaling molecules. Such membrane associations between the mitochondria and a specialized site of the ER, the mitochondria associated-membrane (MAM), as well as between the ER and the plasma membrane (PAM) have been partially characterized at the molecular level. However, historical and recent observations imply that other organelles like peroxisomes, lysosomes, and lipid droplets might also be involved in the formation of such apposing membrane contact sites. Alternatively, reports on so-called mitochondria derived-vesicles (MDV) suggest alternative mechanisms of organelle interaction. Moreover, maintenance of cellular homeostasis requires the precise removal of aged organelles by autophagy-a process which involves the detection of ubiquitinated organelle proteins by the autophagosome membrane, representing another site of membrane associated-signaling. This review will summarize the available data on the existence and composition of organelle contact sites and the molecular specializations each site uses in order to provide a timely overview on the potential functions of organelle interaction.
Keywords: MAM; PAM; endoplasmic reticulum; intracellular signaling; membrane contact sites; mitochondria; organelle dynamics; peroxisomes.
Figures
References
-
- Baerends R. J., Rasmussen S. W., Hilbrands R. E., Van Der Heide M., Faber K. N., Reuvekamp P. T., et al. . (1996). The Hansenula polymorpha PER9 gene encodes a peroxisomal membrane protein essential for peroxisome assembly and integrity. J. Biol. Chem. 271, 8887–8894. 10.1074/jbc.271.15.8887 - DOI - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous