

Detoxification of hostplant's chemical defence rather than its anti-predator co-option drives β-glucosidase-mediated lepidopteran counteradaptation
- PMID: 26443324
- PMCID: PMC4633822
- DOI: 10.1038/ncomms9525
Detoxification of hostplant's chemical defence rather than its anti-predator co-option drives β-glucosidase-mediated lepidopteran counteradaptation
Abstract
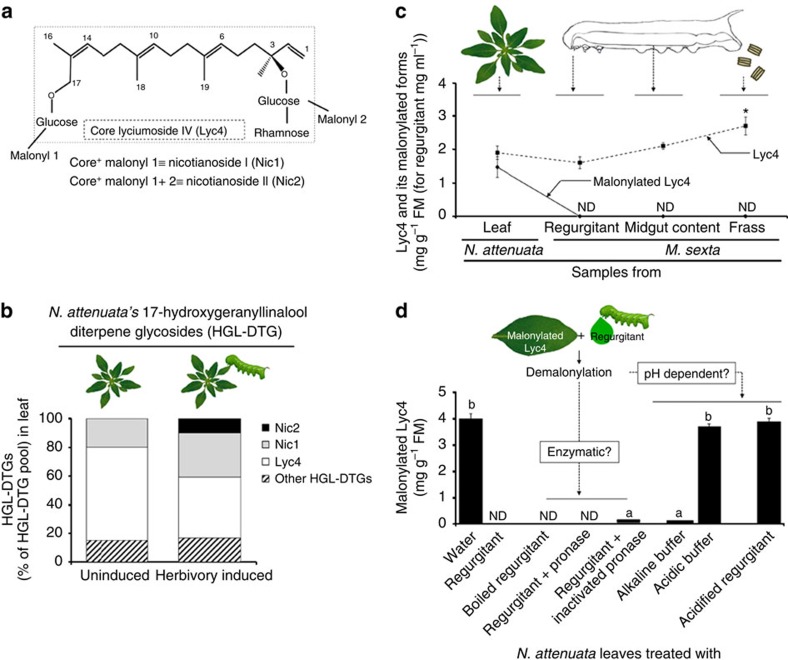
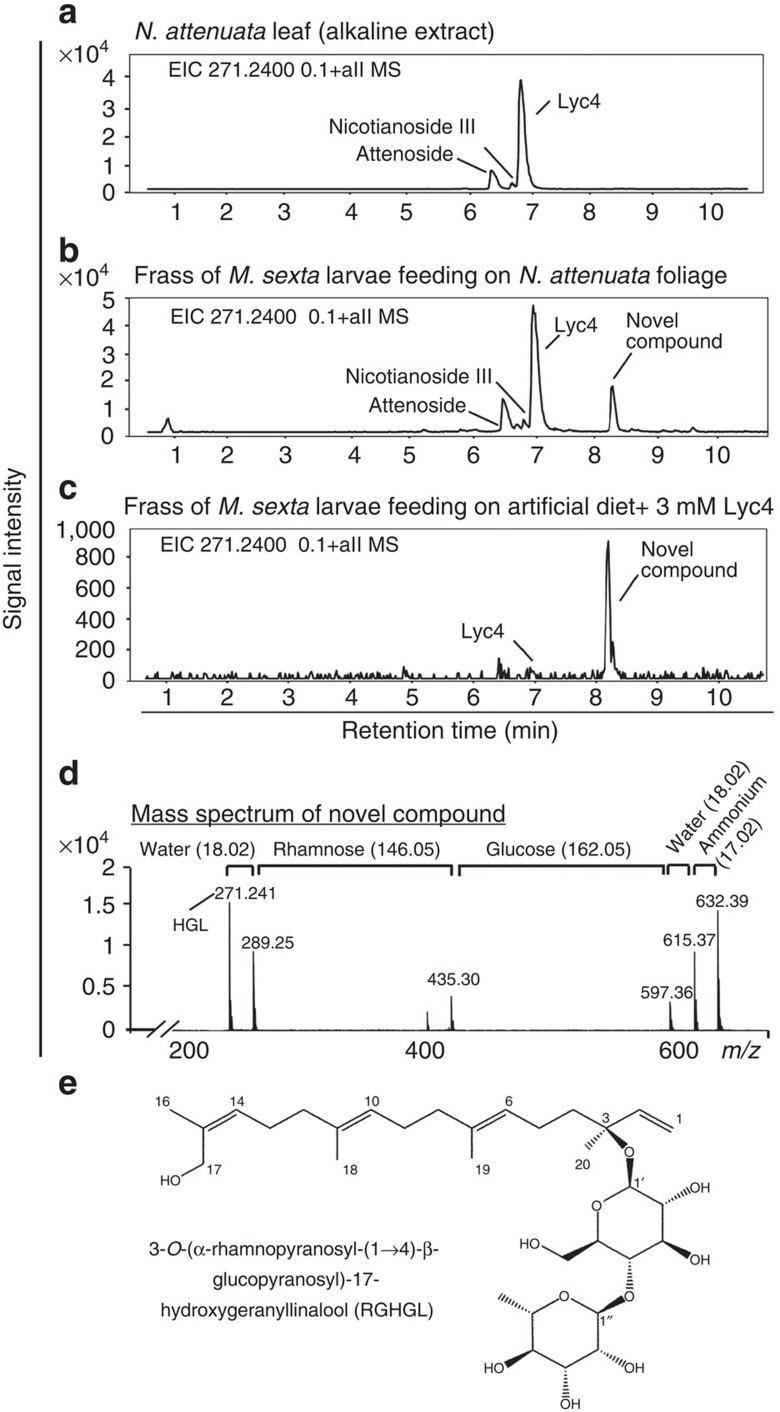
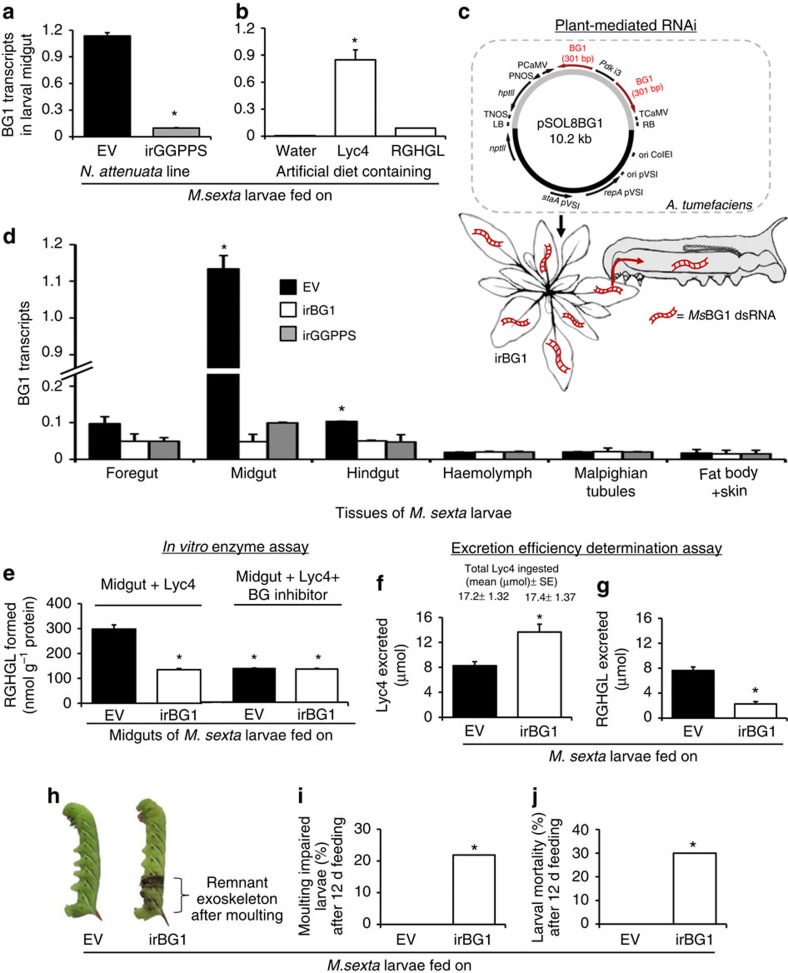
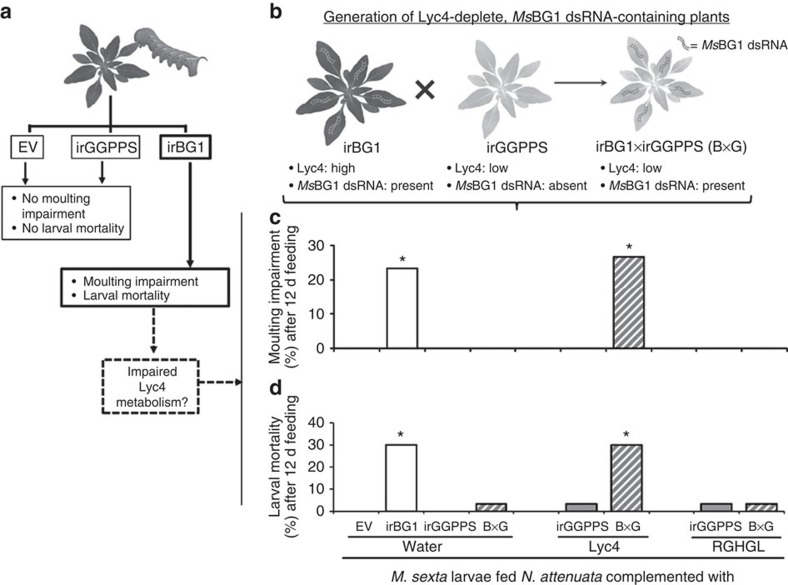
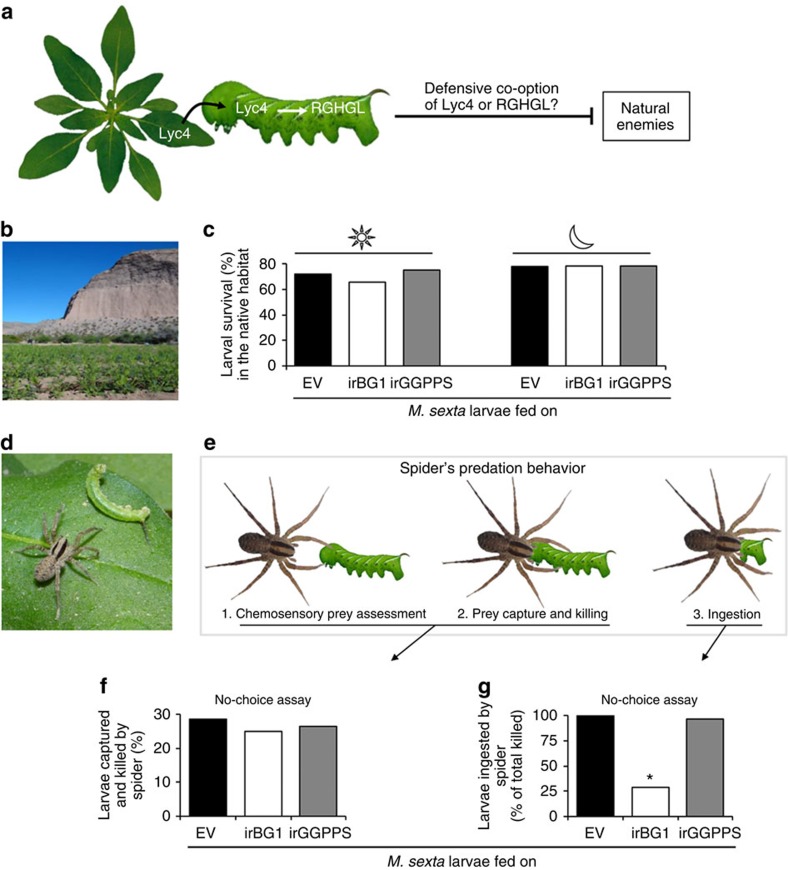
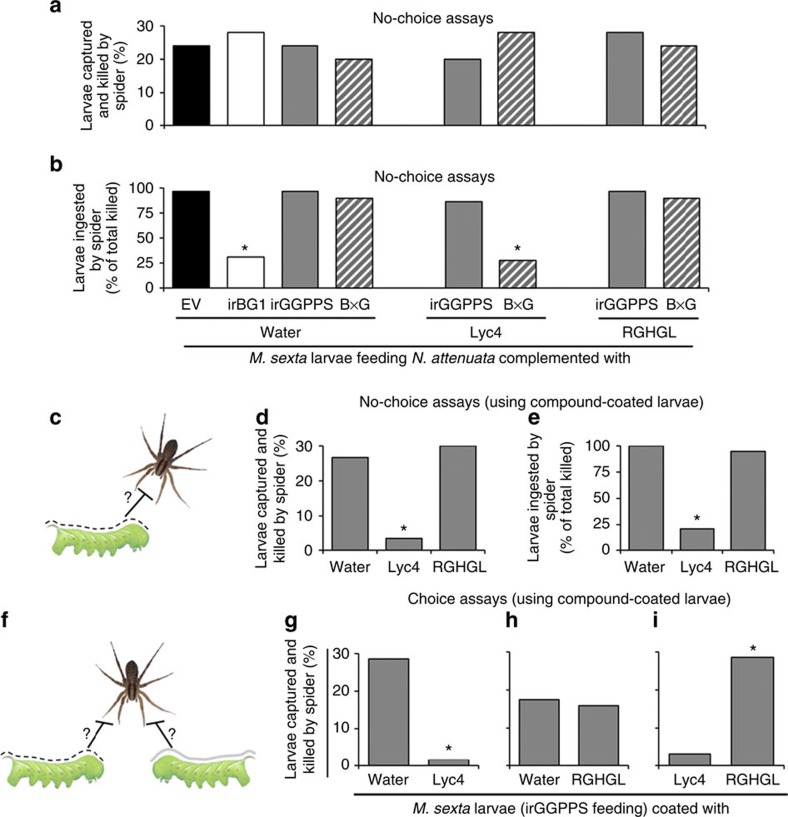
The evolutionary plant-herbivore arms race sometimes gives rise to remarkably unique adaptation strategies. Here we report one such strategy in the lepidopteran herbivore Manduca sexta against its hostplant Nicotiana attenuata's major phytotoxins, 17-hydroxygeranyllinalool diterpene glycoside, lyciumoside IV and its malonylated forms. We show that alkalinity of larval regurgitant non-enzymatically demalonylates the malonylated forms to lyciumoside IV. Lyciumoside IV is then detoxified in the midgut by β-glucosidase 1-catalysed deglycosylation, which is unusual, as typically the deglycosylation of glycosylated phytochemicals by insects results in the opposite: toxin activation. Suppression of deglucosylation by silencing larval β-glucosidase 1 by plant-mediated RNAi causes moulting impairments and mortality. In the native habitat of N. attenuata, β-glucosidase 1 silencing also increases larval unpalatability to native predatory spiders, suggesting that the defensive co-option of lyciumoside IV may be ecologically advantageous. We infer that M. sexta detoxifies this allelochemical to avoid its deleterious effects, rather than co-opting it against predators.
Figures
References
-
- Jones P. & Vogt T. Glycosyltransferases in secondary plant metabolism: tranquilizers and stimulant controllers. Planta 213, 164–174 (2001) . - PubMed
-
- Bowles D., Lim E. K., Poppenberger B. & Vaistij F. E. Glycosyltransferases of lipophilic small molecules. Annu. Rev. Plant Biol. 57, 567–597 (2006) . - PubMed
-
- Pentzold S., Zagrobelny M., Rook F. & Bak S. How insects overcome two-component plant chemical defence: plant β-glucosidases as the main target for herbivore adaptation. Biol. Rev. Camb. Philos. Soc. 89, 531–551 (2014) . - PubMed
-
- Yu S. J. β-Glucosidase in four phytophagous Lepidoptera. Insect Biochem. 19, 103–108 (1989) .
-
- Wouters F. C. et al.. Reglucosylation of the benzoxazinoid DIMBOA with inversion of stereochemical configuration is a detoxification strategy in lepidopteran herbivores. Angew. Chem. Int. Ed. Engl. 53, 11320–11324 (2014) . - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
