

Leukotriene B4 Enhances NOD2-Dependent Innate Response against Influenza Virus Infection
- PMID: 26444420
- PMCID: PMC4596707
- DOI: 10.1371/journal.pone.0139856
Leukotriene B4 Enhances NOD2-Dependent Innate Response against Influenza Virus Infection
Abstract
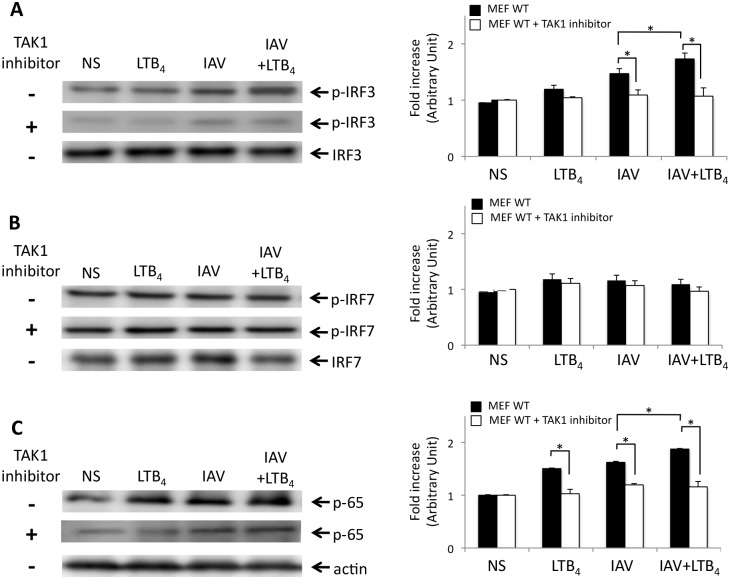
Leukotriene B4 (LTB4), a central mediator of inflammation, is well known for its chemoattractant properties on effectors cells of the immune system. LTB4 also has the ability to control microbial infection by improving host innate defenses through the release of antimicrobial peptides and modulation of intracellular Toll-like receptors (TLRs) expression in response to agonist challenge. In this report, we provide evidences that LTB4 acts on nucleotide-binging oligomerization domain 2 (NOD2) pathway to enhance immune response against influenza A infection. Infected mice receiving LTB4 show improved survival, lung architecture and reduced lung viral loads as compared to placebo-treated animals. NOD2 and its downstream adaptor protein IPS-1 have been found to be essential for LTB4-mediated effects against IAV infection, as absence of NOD2 or IPS-1 diminished its capacity to control viral infection. Treatment of IAV-infected mice with LTB4 induces an increased activation of IPS-1-IRF3 axis leading to an enhanced production of IFNβ in lungs of infected mice. LTB4 also has the ability to act on the RICK-NF-κB axis since administration of LTB4 to mice challenged with MDP markedly increases the secretion of IL-6 and TNFα in lungs of mice. TAK1 appears to be essential to the action of LTB4 on NOD2 pathway since pretreatment of MEFs with TAK1 inhibitor prior stimulation with IAV or MDP strongly abrogated the potentiating effects of LTB4 on both IFNβ and cytokine secretion. Together, our results demonstrate that LTB4, through its ability to activate TAK1, potentiates both IPS-1 and RICK axis of the NOD2 pathway to improve host innate responses.
Conflict of interest statement
Figures







References
-
- Shin EH, Lee HY, Bae YS (2006) Leukotriene B4 stimulates human monocyte-derived dendritic cell chemotaxis. Biochem Biophys Res Commun 348: 606–611. - PubMed
-
- Tager AM, Bromley SK, Medoff BD, Islam SA, Bercury SD, et al. (2003) Leukotriene B4 receptor BLT1 mediates early effector T cell recruitment. Nat Immunol 4: 982–990. - PubMed
-
- Goodarzi K, Goodarzi M, Tager AM, Luster AD, von Andrian UH (2003) Leukotriene B4 and BLT1 control cytotoxic effector T cell recruitment to inflamed tissues. Nat Immunol 4: 965–973. - PubMed
-
- Gaudreault E, Gosselin J (2007) Leukotriene B4-mediated release of antimicrobial peptides against cytomegalovirus is BLT1 dependent. Viral Immunol 20: 407–420. - PubMed
-
- Gosselin J, Borgeat P, Flamand L (2005) Leukotriene B4 protects latently infected mice against murine cytomegalovirus reactivation following allogeneic transplantation. J Immunol 174: 1587–1593. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
