

Distinct Escape Pathway by Hepatitis C Virus Genotype 1a from a Dominant CD8+ T Cell Response by Selection of Altered Epitope Processing
- PMID: 26446603
- PMCID: PMC4702586
- DOI: 10.1128/JVI.01993-15
Distinct Escape Pathway by Hepatitis C Virus Genotype 1a from a Dominant CD8+ T Cell Response by Selection of Altered Epitope Processing
Abstract
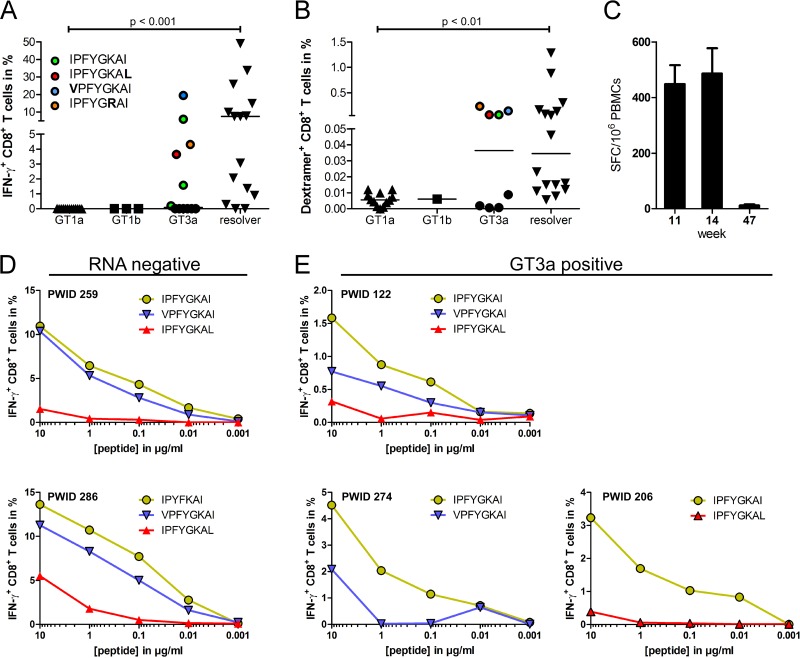
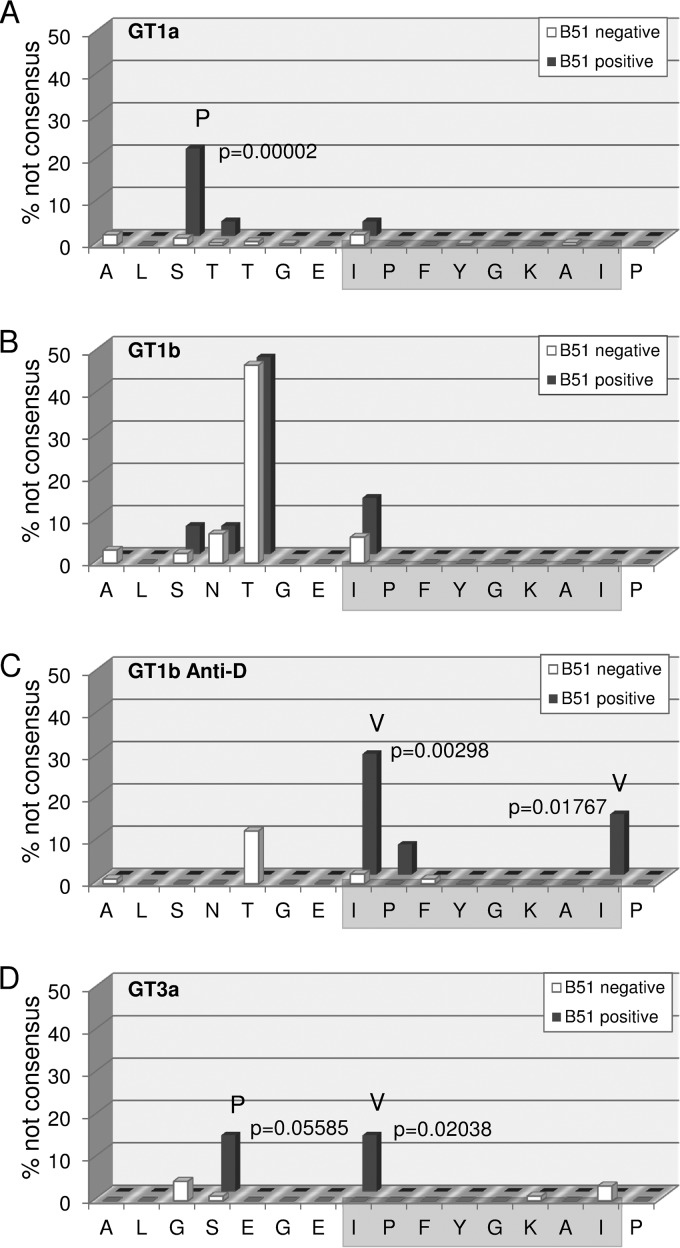
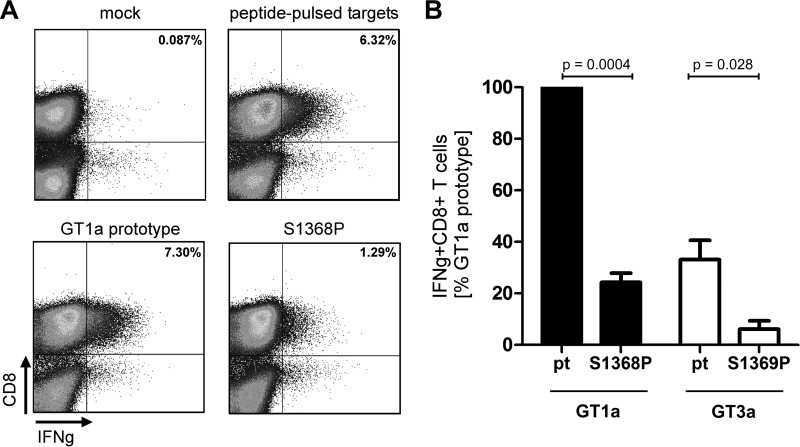
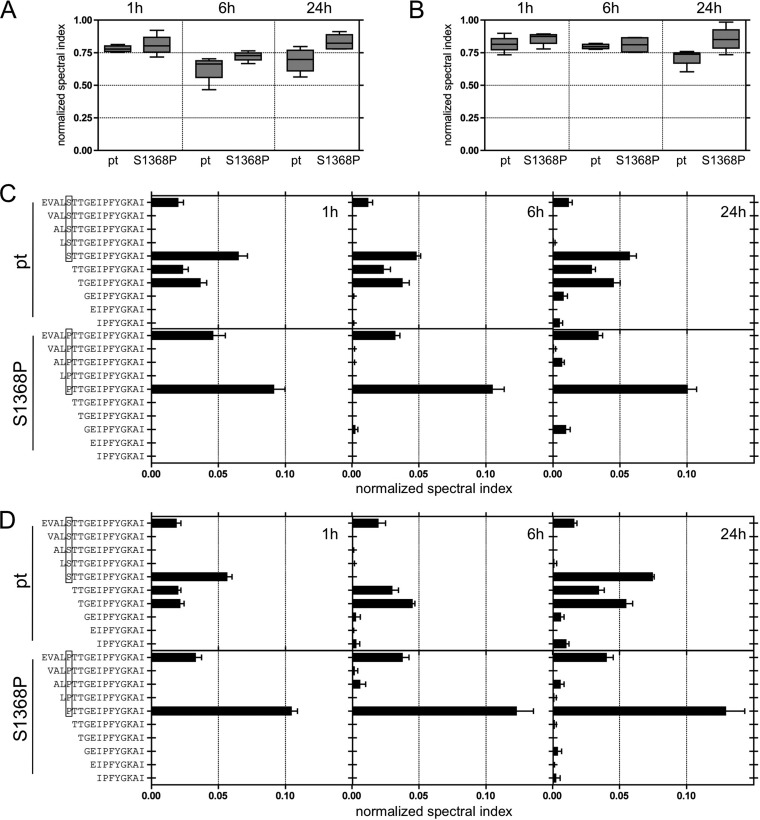
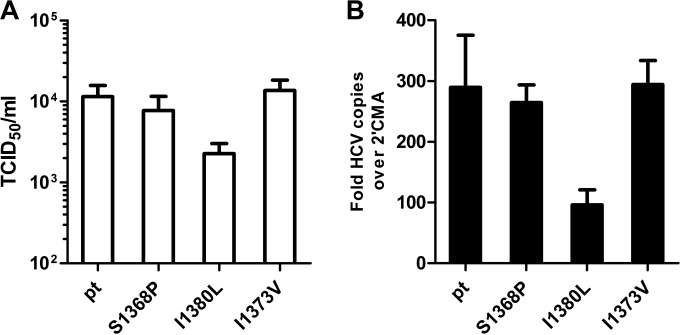
Antiviral CD8(+) T cells are a key component of the adaptive immune response against HCV, but their impact on viral control is influenced by preexisting viral variants in important target epitopes and the development of viral escape mutations. Immunodominant epitopes highly conserved across genotypes therefore are attractive for T cell based prophylactic vaccines. Here, we characterized the CD8(+) T cell response against the highly conserved HLA-B*51-restricted epitope IPFYGKAI1373-1380 located in the helicase domain of NS3 in people who inject drugs (PWID) exposed predominantly to HCV genotypes 1a and 3a. Despite this epitope being conserved in both genotypes, the corresponding CD8(+) T cell response was detected only in PWID infected with genotype 3a and HCV-RNA negative PWID, but not in PWID infected with genotype 1a. In genotype 3a, the detection of strong CD8(+) T cell responses was associated with epitope variants in the autologous virus consistent with immune escape. Analysis of viral sequences from multiple cohorts confirmed HLA-B*51-associated escape mutations inside the epitope in genotype 3a, but not in genotype 1a. Here, a distinct substitution in the N-terminal flanking region located 5 residues upstream of the epitope (S1368P; P = 0.00002) was selected in HLA-B*51-positive individuals. Functional assays revealed that the S1368P substitution impaired recognition of target cells presenting the endogenously processed epitope. The results highlight that, despite an epitope being highly conserved between two genotypes, there are major differences in the selected viral escape pathways and the corresponding T cell responses.
Importance: HCV is able to evolutionary adapt to CD8(+) T cell immune pressure in multiple ways. Beyond selection of mutations inside targeted epitopes, this study demonstrates that HCV inhibits epitope processing by modification of the epitope flanking region under T cell immune pressure. Selection of a substitution five amino acids upstream of the epitope underlines that efficient antigen presentation strongly depends on its larger sequence context and that blocking of the multistep process of antigen processing by mutation is exploited also by HCV. The pathways to mutational escape of HCV are to some extent predictable but are distinct in different genotypes. Importantly, the selected escape pathway of HCV may have consequences for the destiny of antigen-specific CD8(+) T cells.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
References
-
- Page K, Hahn JA, Evans J, Shiboski S, Lum P, Delwart E, Tobler L, Andrews W, Avanesyan L, Cooper S, Busch MP. 2009. Acute hepatitis C virus infection in young adult injection drug users: a prospective study of incident infection, resolution, and reinfection. J Infect Dis 200:1216–1226. doi:10.1086/605947. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
