

doi: 10.1128/JVI.01671-15.
Print 2016 Jan 1.
Mismatches in the Influenza A Virus RNA Panhandle Prevent Retinoic Acid-Inducible Gene I (RIG-I) Sensing by Impairing RNA/RIG-I Complex Formation
Affiliations
- PMID: 26446607
- PMCID: PMC4702558
- DOI: 10.1128/JVI.01671-15
Item in Clipboard
Mismatches in the Influenza A Virus RNA Panhandle Prevent Retinoic Acid-Inducible Gene I (RIG-I) Sensing by Impairing RNA/RIG-I Complex Formation
J Virol.
.
Abstract
Influenza virus RNA (vRNA) promoter panhandle structures are believed to be sensed by retinoic acid-inducible gene I (RIG-I). The occurrence of mismatches in this double-stranded RNA structure raises questions about their effect on innate sensing. Our results suggest that mismatches in vRNA promoters decrease binding to RIG-I in vivo, affecting RNA/RIG-I complex formation and preventing RIG-I activation. These results can be inferred to apply to other viruses and suggest that mismatches may represent a general viral strategy to escape RIG-I sensing.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
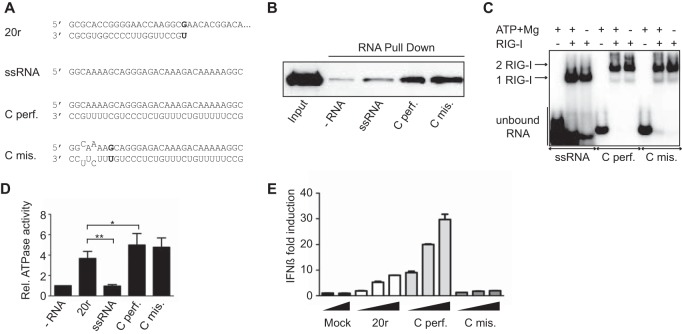
Mismatches do not affect RIG-I binding and ATPase activity but prevent IFN-β activation. (A) Sequences of the RNAs used to mimic the influenza virus cRNA panhandle structures with (mis.) or without (perf.) mismatches. (B) RIG-I binding. RNA pulldown assays were performed with 13 pmol of biotinylated RNA ligands (as previously described [12]) and 13 pmol of His–RIG-I (16). Reaction products were analyzed by Western blotting with antihistidine antibody (1:2,000, H1029; Sigma). (C) RIG-I oligomerization was monitored by EMSA. Radiolabeled RNAs (25 pmol) were incubated with purified His–RIG-I (50 pmol of RIG-I). Reaction products were analyzed on native gradient acrylamide gel and revealed by phosphorimaging (Typhoon; GE Healthcare Life Sciences) (12). (D) Relative (Rel.) RIG-I ATPase activity. Purified RIG-I was incubated at 37°C with [γ-32P]ATP in the presence of various RNA ligands as indicated. Data are presented as the mean ± the standard error of the mean (n = 4). Statistical significance: NS, P > 0.05; *, 0.01 ≤ P < 0.05; **, P < 0.001 (12). (E) IFN-β promoter activation. A549 cells were transfected with luciferase-based reporter gene plasmids (pβ-IFN-fl-lucter, which carries the firefly luciferase gene driven by the human IFN-β promoter [17], and pTK-rl-lucter, which carries the Renilla luciferase-encoding gene [Promega] driven by the herpes simplex virus thymidine kinase promoter) and increasing amounts (200, 400, and 800 ng) of the RNAs indicated. Relative luciferase activation (fold IFN-β induction) was calculated by normalization to the mock-treated control and is represented as the mean ± the standard deviation (n = 2) (12).
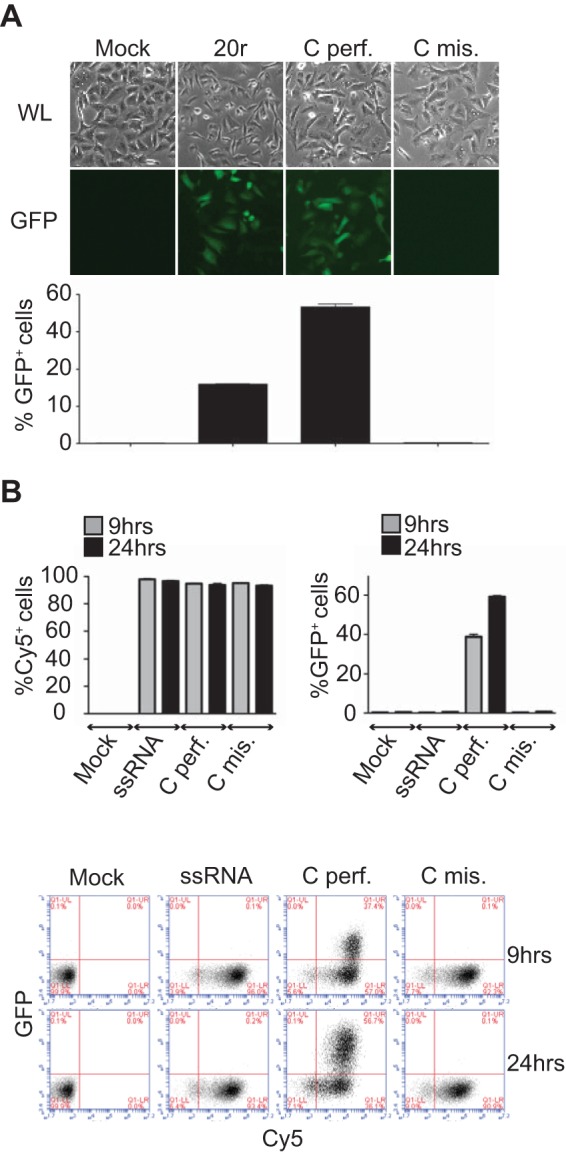
Mismatches do not affect RNA stability. (A, B) IFN-β promoter activation. A549/pr(IFN-β).GFP reporter cells (13) were transfected with 400 ng of the RNAs indicated for 24 h before fluorescence-activated cell sorter (FACS) analysis. (A) At the top are white light (WL) and fluorescence (GFP) microscope pictures. At the bottom are flow cytometry data. Results are plotted as percentages of GFP-positive cells (x axis). Data are presented as means ± standard deviations (n = 2). (B) RNA stability. A549/pr(IFN-β).GFP reporter cells were transfected with Cy5-labeled RNAs, and Cy5 fluorescence was monitored over time by FACS analysis. Results are plotted as percentages of Cy5- or GFP-positive cells. Data are presented as means ± standard deviations (n = 2). At the bottom are representative plots of the FACS data. Results are plotted as percentages of Cy5 (x axis)- and GFP (y axis)-positive cells.
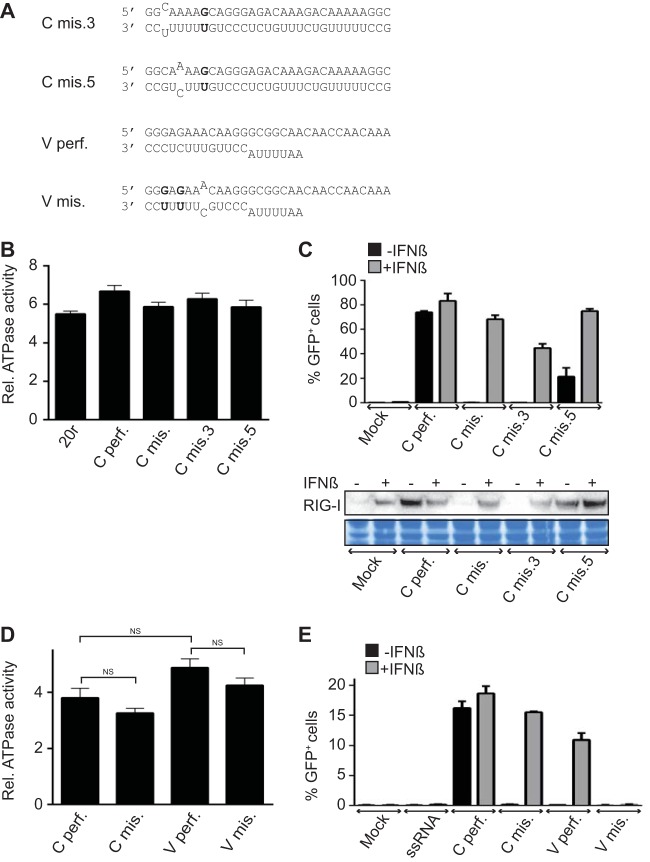
A single mismatch on dsRNA is enough to limit IFN-β induction. (A) Sequences of the RNAs used. (B) RIG-I ATPase activity. Purified RIG-I was incubated at 37°C with [γ-32P]ATP in the presence of various RNA ligands as indicated. Data are presented as the mean ± the standard error of the mean (n = 4). Statistical significance: NS, P > 0.05; *, 0.01 ≤ P < 0.05; **, P < 0.001 (12). (C) IFN-β promoter activation. A549/pr(IFN-β).GFP reporter cells were transfected with 500 ng of the RNAs indicated for 24 h with or without IFN-β treatment 6 h prior to transfection. At the top are the percentages of GFP-positive cells measured by FACS analysis. Data are presented as the mean ± the standard deviation (n = 2). At the bottom are the intracellular levels of RIG-I analyzed by Western blotting. As a loading control, the membrane was stained with Coomassie blue after immunoblotting. (D) RNA-induced ATPase activity of RIG-I. Purified RIG-I was incubated at room temperature with [γ-32P]ATP in the presence of various RNA ligands as indicated. Data are presented as the mean ± the standard error of the mean (n = 5). Statistical significance: NS, P > 0.05; **, P < 0.001. (E) IFN-β promoter activation. A549/pr(IFN-β).GFP reporter cells were transfected with 500 ng of the RNAs indicated for 24 h with or without IFN-β treatment 6 h prior to transfection. Results are plotted as the percentages of GFP-positive cells measured by flow cytometry. Data are presented as the mean ± the standard deviation (n = 2).
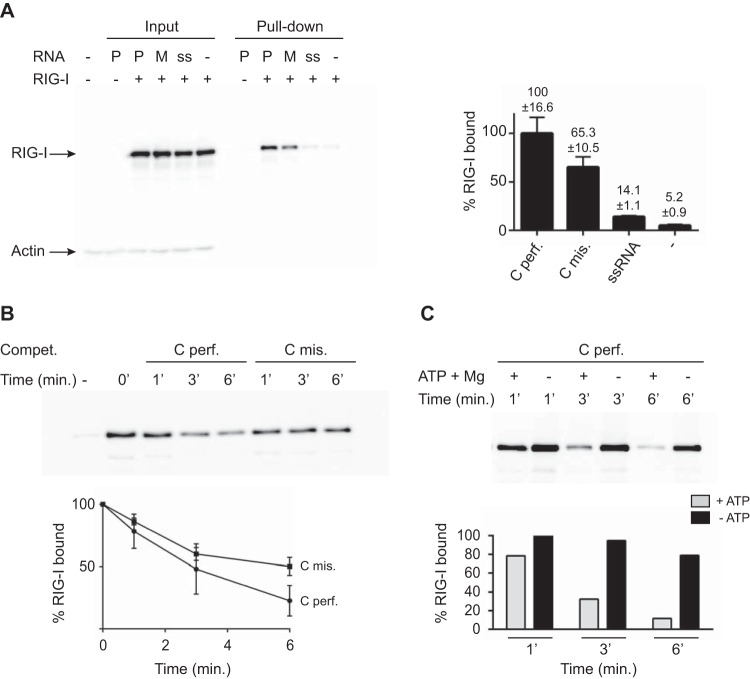
Mismatches destabilize RNA/RIG-I complex formation. (A) In vivo RIG-I binding. 293T cells were first transfected with pEF-Bos-RIG-I (2 μg) for 24 h and then transfected with 2 μg of biotinylated perfect cRNA (Cperf.) or mismatch RNA (Cmis.). As a control, cells were transfected with 2 μg of biotinylated ssRNA. At 2 h after RNA transfection, cells were lysed and lysates were incubated with streptavidin beads for 2 h at 4°C to pull down RNA-bound RIG-I. Reactions were analyzed by Western blotting with anti-RIG-I antibody (ALX-804-849-C100; Enzo Life Sciences). Data are presented as the mean ± the standard deviation (n = 2). (B) Competition (Compet.) in an RNA pulldown assay. Thirteen picomoles of biotinylated perfect cRNA was bound to streptavidin beads by incubation for 2 h at 4°C. Thirteen picomoles of His–RIG-I protein was added to the reaction mixtures for 15 min of incubation at 37°C. A competition assay was performed with a 3-fold excess of either nonbiotinylated perfect (Cperf.) or mismatched (Cmis.) cRNA for 1, 3, or 6 min at 37°C in the presence of ATP and MgCl2. RIG-I binding was monitored by Western blotting and quantified with anti-RIG-I antibody. After quantification, results were plotted as the percentage of RIG-I that remained bound to the beads as a function of time. Data are presented as the mean ± the standard deviation (n = 5). (C) Competition in an RNA pulldown assay. Thirteen picomoles of biotinylated perfect cRNA was bound to streptavidin beads by incubation for 2 h at 4°C. Thirteen picomoles of His–RIG-I protein was added to the reaction mixtures for 15 min of incubation at 37°C. A competition assay was performed with a 3-fold excess of either nonbiotinylated perfect cRNA (Cperf.) for 1, 3, or 6 min at 37°C in the absence or presence of ATP and MgCl2 as indicated. RIG-I binding was monitored by Western blotting and quantified with anti-RIG-I antibody. A representative experiment is shown. Western blot quantifications were performed with ImageJ version 1.44p (National Institutes of Health, Bethesda, MD; http://imagej.nih.gov/ij/ ).
References
-
- Goubau D, Schlee M, Deddouche S, Pruijssers AJ, Zillinger T, Goldeck M, Schuberth C, Van der Veen AG, Fujimura T, Rehwinkel J, Iskarpatyoti JA, Barchet W, Ludwig J, Dermody TS, Hartmann G, Reis e Sousa C. 2014. Antiviral immunity via RIG-I-mediated recognition of RNA bearing 5′-diphosphates. Nature 514:372–375. doi:10.1038/nature13590. - DOI - PMC - PubMed
-
- Weber M, Gawanbacht A, Habjan M, Rang A, Borner C, Schmidt AM, Veitinger S, Jacob R, Devignot S, Kochs G, Garcia-Sastre A, Weber F. 2013. Incoming RNA virus nucleocapsids containing a 5′-triphosphorylated genome activate RIG-I and antiviral signaling. Cell Host Microbe 13:336–346. doi:10.1016/j.chom.2013.01.012. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
