

Genome engineering: Drosophila melanogaster and beyond
- PMID: 26447401
- PMCID: PMC4761275
- DOI: 10.1002/wdev.214
Genome engineering: Drosophila melanogaster and beyond
Abstract
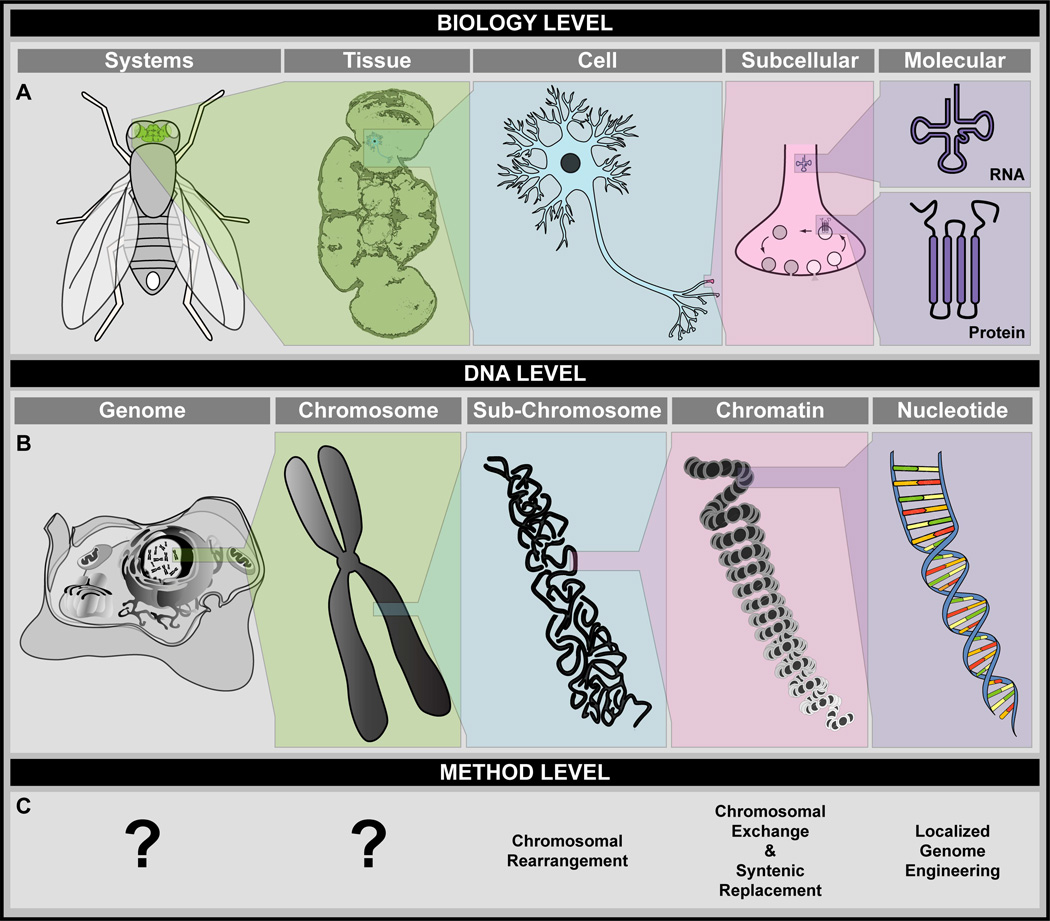
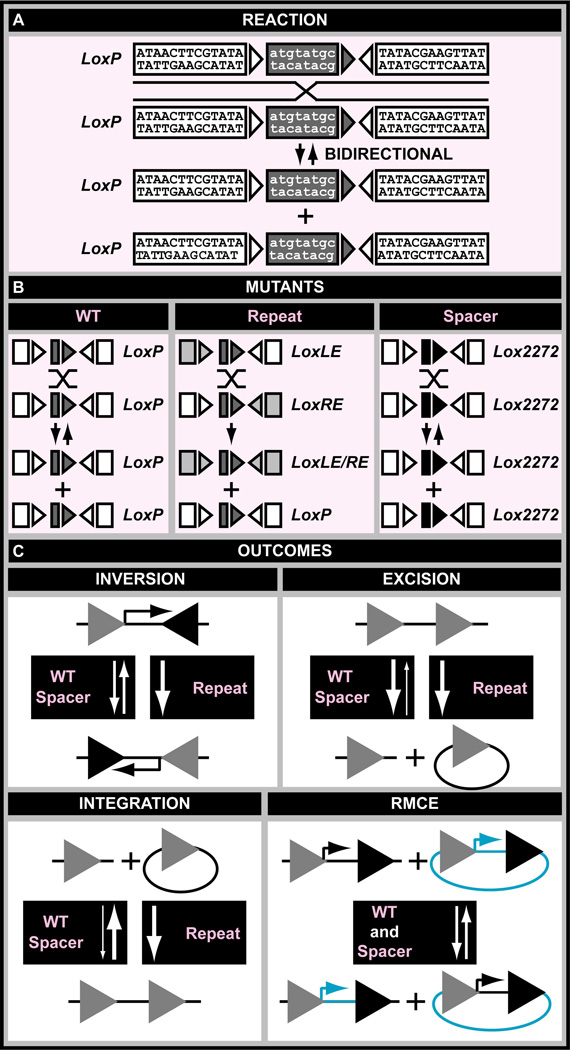
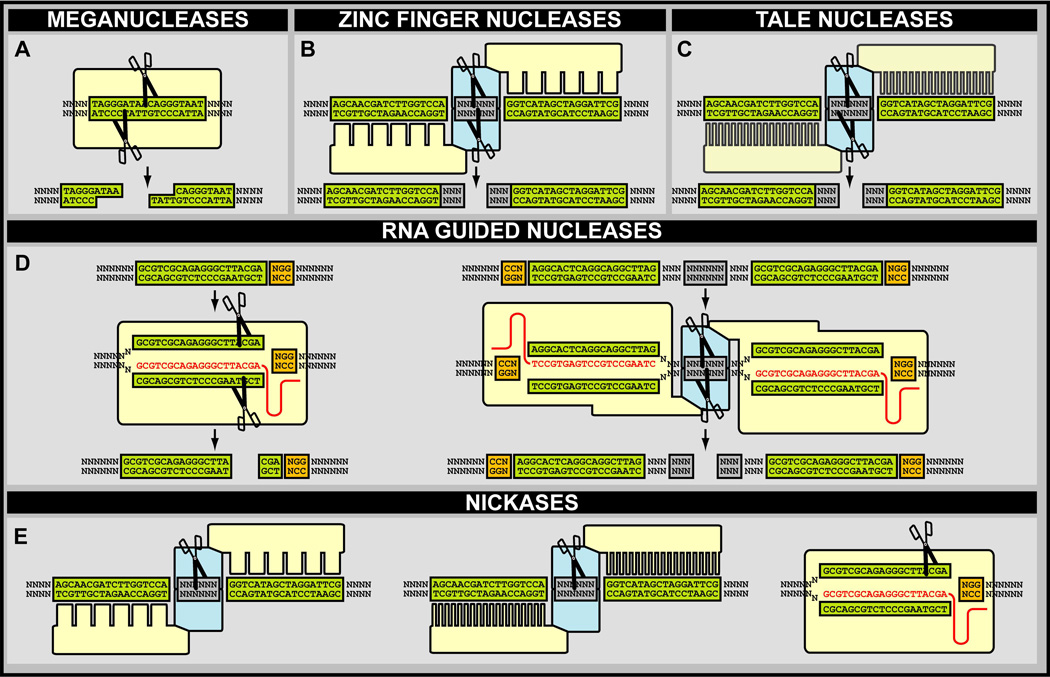
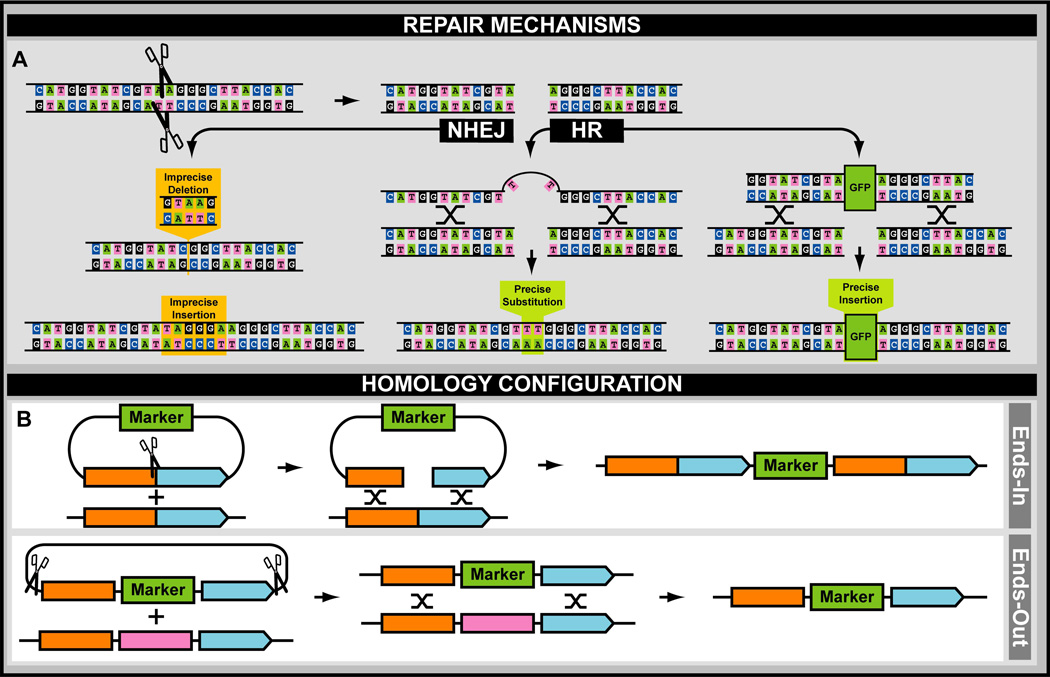
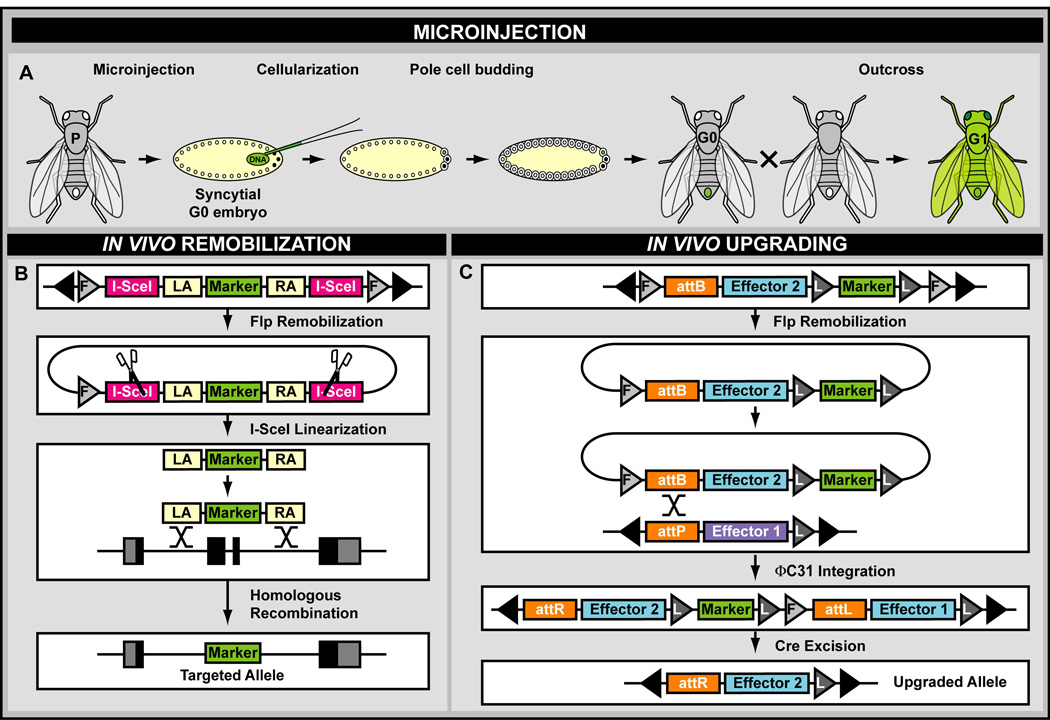
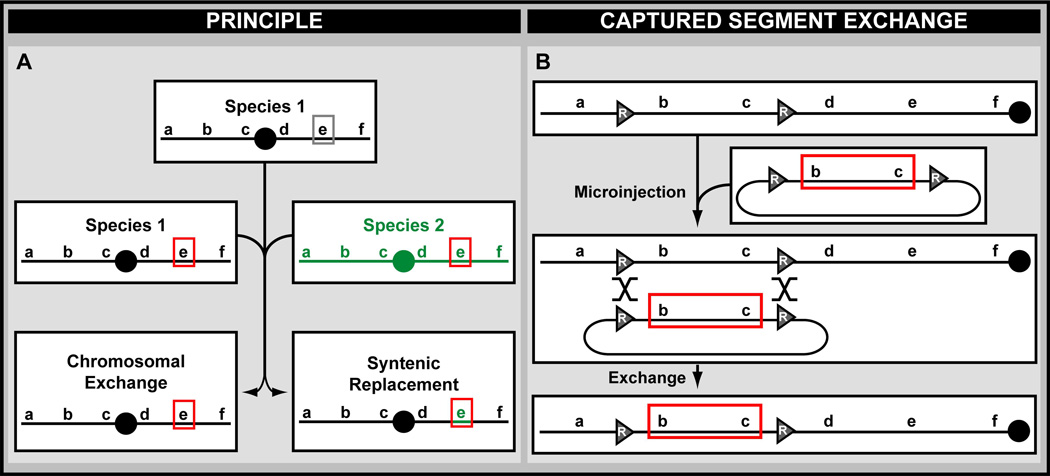
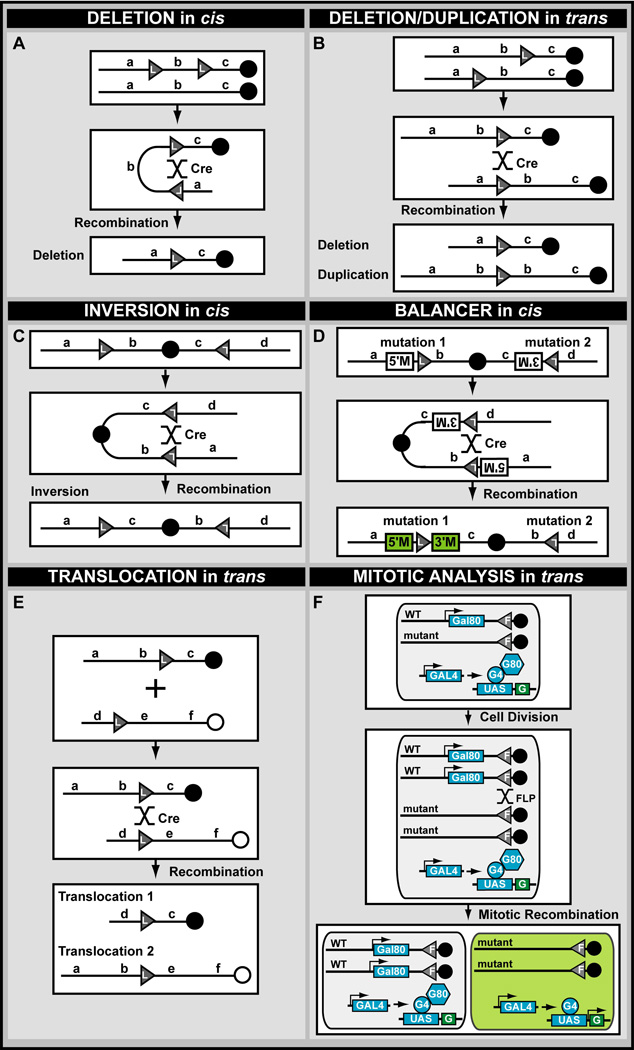
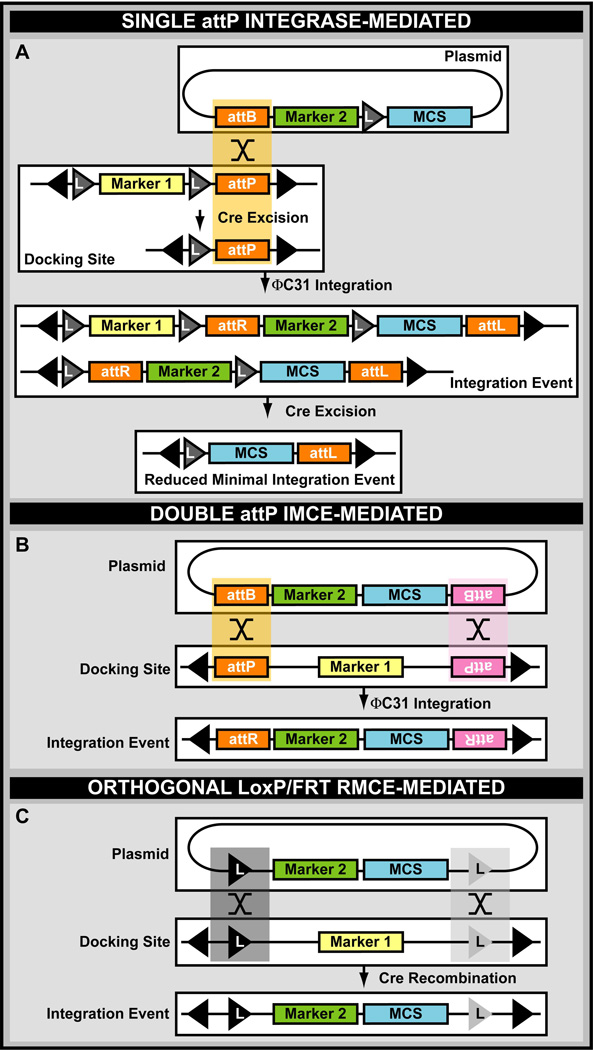
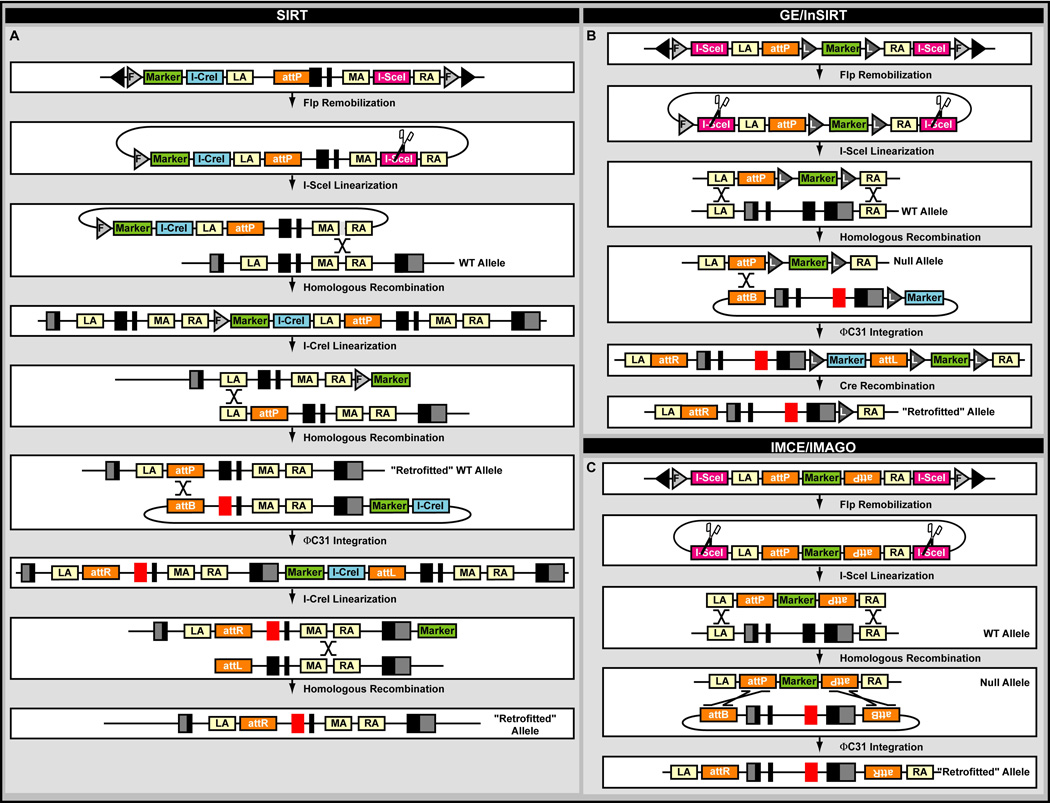
A central challenge in investigating biological phenomena is the development of techniques to modify genomic DNA with nucleotide precision that can be transmitted through the germ line. Recent years have brought a boon in these technologies, now collectively known as genome engineering. Defined genomic manipulations at the nucleotide level enable a variety of reverse engineering paradigms, providing new opportunities to interrogate diverse biological functions. These genetic modifications include controlled removal, insertion, and substitution of genetic fragments, both small and large. Small fragments up to a few kilobases (e.g., single nucleotide mutations, small deletions, or gene tagging at single or multiple gene loci) to large fragments up to megabase resolution can be manipulated at single loci to create deletions, duplications, inversions, or translocations of substantial sections of whole chromosome arms. A specialized substitution of chromosomal portions that presumably are functionally orthologous between different organisms through syntenic replacement, can provide proof of evolutionary conservation between regulatory sequences. Large transgenes containing endogenous or synthetic DNA can be integrated at defined genomic locations, permitting an alternative proof of evolutionary conservation, and sophisticated transgenes can be used to interrogate biological phenomena. Precision engineering can additionally be used to manipulate the genomes of organelles (e.g., mitochondria). Novel genome engineering paradigms are often accelerated in existing, easily genetically tractable model organisms, primarily because these paradigms can be integrated in a rigorous, existing technology foundation. The Drosophila melanogaster fly model is ideal for these types of studies. Due to its small genome size, having just four chromosomes, the vast amount of cutting-edge genetic technologies, and its short life-cycle and inexpensive maintenance requirements, the fly is exceptionally amenable to complex genetic analysis using advanced genome engineering. Thus, highly sophisticated methods developed in the fly model can be used in nearly any sequenced organism. Here, we summarize different ways to perform precise inheritable genome engineering using integrases, recombinases, and DNA nucleases in the D. melanogaster. For further resources related to this article, please visit the WIREs website.
© 2015 Wiley Periodicals, Inc.
Figures


References
-
- Karas BJ, Suzuki Y, Weyman PD. Strategies for cloning and manipulating natural and synthetic chromosomes. Chromosome Res. 2015;23(1573-6849 (Electronic)):57–68. - PubMed
-
- Gibson DG, Benders GA, Andrews-Pfannkoch C, et al. Complete chemical synthesis, assembly, and cloning of a Mycoplasma genitalium genome. Science. 2008;319(5867):1215–1220. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
