

Review
doi: 10.1016/j.neuron.2015.09.017.
From Behavior to Neural Dynamics: An Integrated Theory of Attention
Affiliations
- PMID: 26447577
- PMCID: PMC4604109
- DOI: 10.1016/j.neuron.2015.09.017
Item in Clipboard
Review
From Behavior to Neural Dynamics: An Integrated Theory of Attention
Neuron.
.
Abstract
The brain has a limited capacity and therefore needs mechanisms to selectively enhance the information most relevant to one's current behavior. We refer to these mechanisms as "attention." Attention acts by increasing the strength of selected neural representations and preferentially routing them through the brain's large-scale network. This is a critical component of cognition and therefore has been a central topic in cognitive neuroscience. Here we review a diverse literature that has studied attention at the level of behavior, networks, circuits, and neurons. We then integrate these disparate results into a unified theory of attention.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
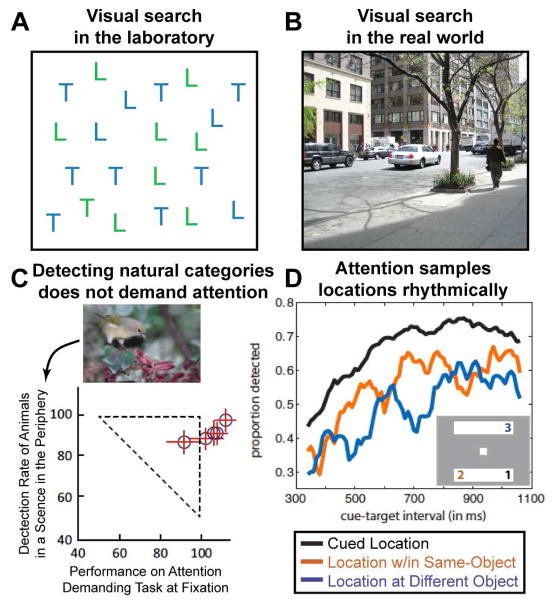
(A) Visual search in artificial displays versus in real-world scenes. Detecting the presence of a green T (conjunction search) is effortful and time-consuming, such that reaction times increase as a function of display items. (B) In contrast, detecting the presence of categorical object information such as ‘people’ or ‘cars’ in real-world scenes requires only a single glance, despite the large number and variety of distracter objects. (C) The detection of animals or vehicles in natural scenes does not require focused spatial attention. In a dual-task paradigm, subjects performed a central discrimination task, while detecting animals in scenes presented in the periphery. Performance is normalized to a condition when only a single task was performed. Performance on the peripheral animal detection task was only mildly impaired by simultaneously performing the central discrimination task. Adapted from (Li et al., 2002). (D) Selective attention has rhythmic properties. Subjects detected the dimming of a part of a rectangular object at a spatially cued (black line; location #1 in the two object display depicted as an example), or at an uncued location of the same object than the cued location (orange line; location #2), or a different object (blue line; location #3). Accuracy is plotted as a function of the cue-target interval revealing the following rhythmic properties: at the cued location, detection performance fluctuated at 8Hz, whereas at the same- and different-object locations a characteristic anti-phase relationship of a 4Hz rhythm was observed. Adapted from (Fiebelkorn et al., 2013).
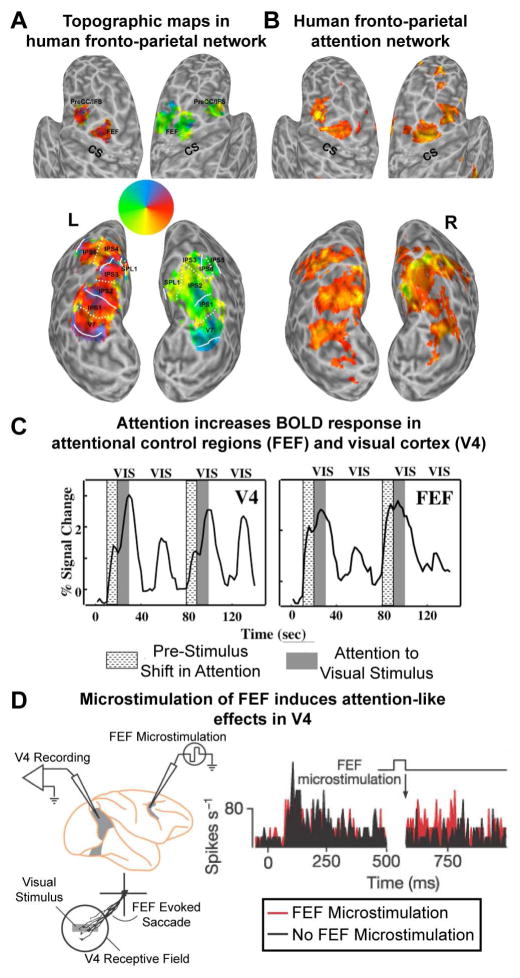
(A) Topographic organization of areas in human frontal and parietal cortex. Using a memory-guided saccade task several areas with a systematic representation of the contralateral visual field were identified along the intraparietal sulcus (IPS0-5), adjacent superior parietal cortex (SPL1), and in superior (FEF) and inferior aspects of precentral cortex. Adapted from (Silver and Kastner, 2009). (B) Attention-related activations within parietal and frontal cortex in a spatial attention task. There is significant overlap between attention-related activations and topographic representations in higher-order cortex. Adapted from (Szczepanski et al., 2010). (C) Time series of fMRI signals in V4 and FEF. Directing attention to a peripheral target location in the absence of visual stimulation led to an increase of baseline activity (textured blocks), which was followed by a further increase after the onset of the stimuli (gray shaded blocks) in V4, but not in FEF, where the initially stronger baseline increase was sustained, thus reflecting the attentional operations of the task and not sensory processing. Adapted from (Kastner et al., 1999) (D) Microstimulation of FEF (below the threshold that evokes an eye movement) induces attention-like increases in the spikes/s of V4 neurons with receptive fields that overlap the FEF site (black is baseline; red with microstimulation). Adapted from (Moore and Armstrong, 2003).
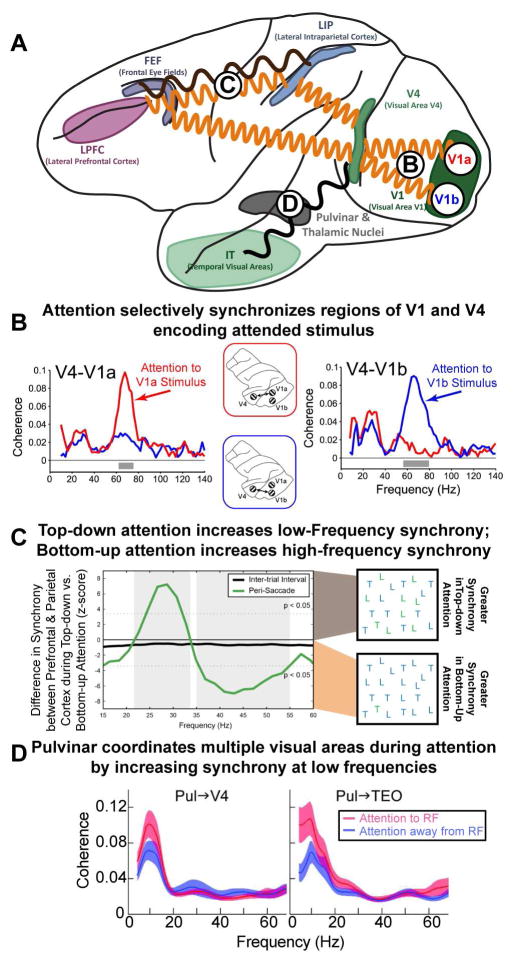
(A) Attention modulates the synchrony between different brain regions. A minor subset of the effects of attention is outlined. Circles with letters refer to other parts of the figure. (B) Attentional modulation is specific to selected objects: synchrony between V4 and V1 is specific to those regions that encode the currently attended stimulus (red for a stimulus encoded by V1a; blue for V1b). Note that these changes in the pattern of synchrony overlay the underlying anatomy and can occur rapidly, with each shift in attention. Adapted from (Bosman et al., 2012). (C) There is also flexibility in the frequency of oscillations between brain regions. Internal (top-down) direction of attention and external (bottom-up) capture of attention emphasize different frequency bands between the same brain regions. Synchrony between prefrontal cortex (PFC) and parietal cortex (PPC) changed frequency depending on the type of attention. When attention was externally captured by a salient stimulus, this was reflected in neural activity in PPC first, followed by PFC. In addition, synchrony between PFC and PPC was observed at high-frequencies (~45 Hz; shown as negative deflection). In contrast, when attention was internally directed by the memory of a target stimulus (as in visual search) neural activity was found first in PFC and then PPC and PFC-PPC synchrony was observed at low frequencies (~25 Hz; shown as positive deflection). Adapted from (Buschman and Miller, 2007) (D) Sub-cortical regions, such as the pulvinar, play an important role in attention. In addition, they may act to coordinate activity across cortical regions: attention increased low-frequency synchrony between the pulvinar and V4/TEO (left/right, respectively), when allocated at the receptive field (RF) represented by the recorded neurons (ATT to RF) relative to a different RF location (ATT away from RF). This may organize higher-frequency oscillations, facilitating the establishment of synchrony observed between cortical regions (as in B and C). Adapted from (Saalmann et al., 2012).
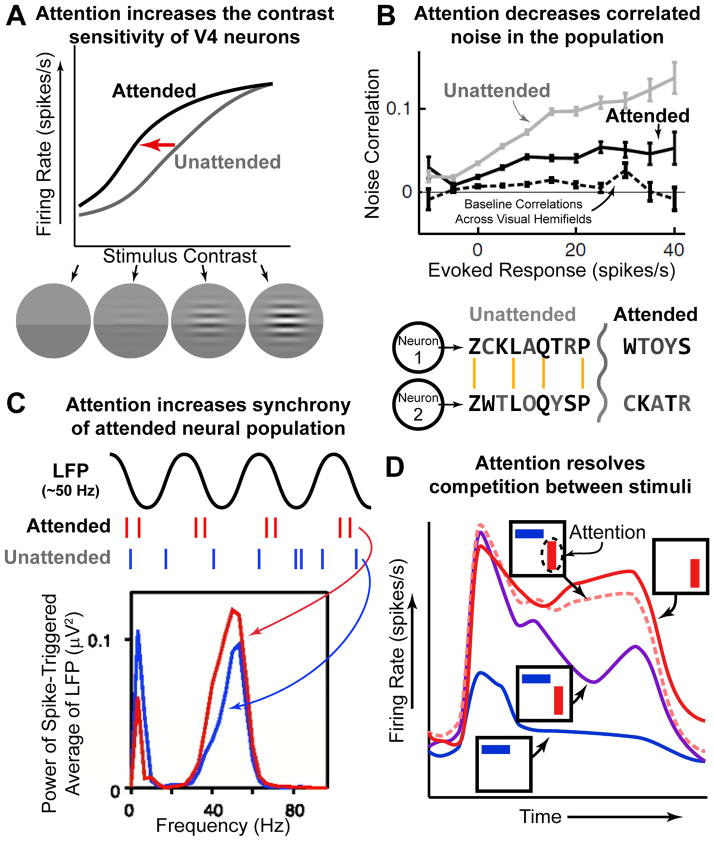
(A) Attention increases the sensitivity of V4 neurons. Neurons increase their firing rate response to stimuli of increasing contrast; even without attention (gray line). Attending to the stimulus increases the response to stimuli at lower contrasts (black line; note leftward shift). Data is schematized from (Reynolds et al., 2000). (B) Attention reduces noise correlations in neural activity. Simultaneously recorded neurons often share uninformative ‘noise’ signals. This is schematized in the two model neurons below. Noise is schematized as black letters in the encode ‘stream’, gray letters are ‘signal’. The two neurons share much of the same ‘noise’; reducing such noise makes the message encoded by both neurons clearer (‘toy car’). Attention has such an effect in the brain (top figure; black line is below gray line). Adapted from (Cohen and Maunsell, 2009). (C) Attention increases the synchrony of selected neurons. The synchrony between individual neurons and the population (as measured by the local field potential, LFP) increases with attention allocated at RF (‘attended’, red) relative to away from RF (‘unattended’, blue). This effect is specific to a ‘gamma’ oscillation at ~50 Hz (note that synchrony is reduced at low-frequency, ~10 Hz, oscillations). Adapted from (Fries et al, 2001) (D) Attention resolves competition between stimuli. Stimuli compete for representation in V4 neurons: the response to two stimuli (purple) is approximately the average of the response to either stimuli when presented alone (red, preferred, and blue, non-preferred). Attending to a single stimulus ‘rescues’ this competition, causing the neuron to respond as if only the attended stimulus was presented (pink dashed line). Data is schematized from (Reynolds et al., 1999).
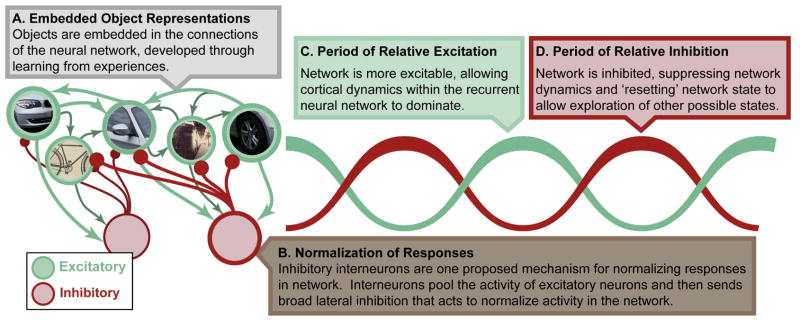
We propose many of the effects of attention to be due to local interactions within a cortical region. Attentional selection interacts with bottom-up sensory drive (not shown) as well as object representations that have been embedded within the neural network through learning (A). Attention acts on these representations by changing interactions between excitatory neurons (green) and local inhibitory interneurons (red). In particular, pooling of responses by inhibitory interneurons could form the basis for normalization of excitatory responses (B). As noted in the main text, normalization likely plays a key role in perception and attention. Furthermore, rhythmic interactions between excitatory and inhibitory neurons is proposed to underlie high-frequency oscillations (C), which are increased with attention. These oscillations may play a fundamental role to temporally organize neural activity. For example, periods of inhibition may ‘reset’ the network, allowing it to explore more than one attractor state (details in main text).
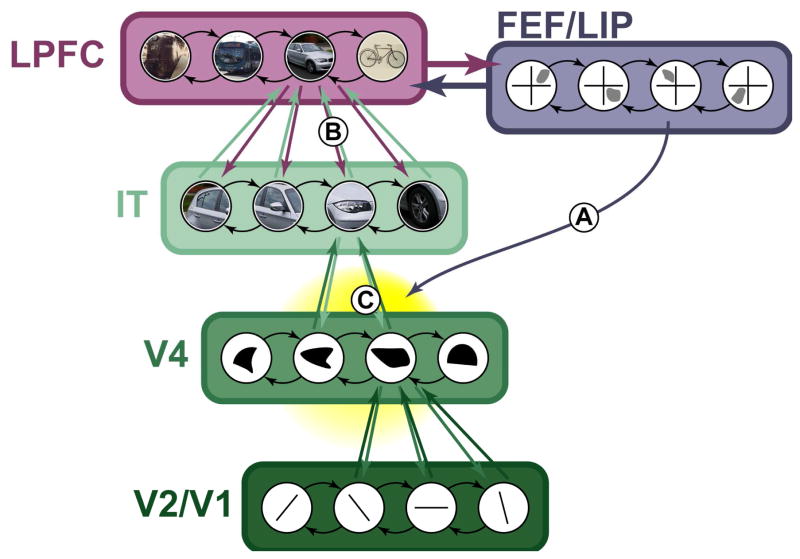
The brain is a densely interconnected network and so attentional selections, whether (A) spatial or (B) featural, propagate up and down the visual hierarchy. In this way, they will interact (C), allowing spatial attention to lead to selection of objects with similar features or featural selection to drive spatial attention.
References
-
- Awh E, Jonides J. Overlapping mechanisms of attention and spatial working memory. Trends Cogn Sci. 2001;5:119–126. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
