

Utilization of Glycosaminoglycans/Proteoglycans as Carriers for Targeted Therapy Delivery
- PMID: 26448753
- PMCID: PMC4581573
- DOI: 10.1155/2015/537560
Utilization of Glycosaminoglycans/Proteoglycans as Carriers for Targeted Therapy Delivery
Abstract
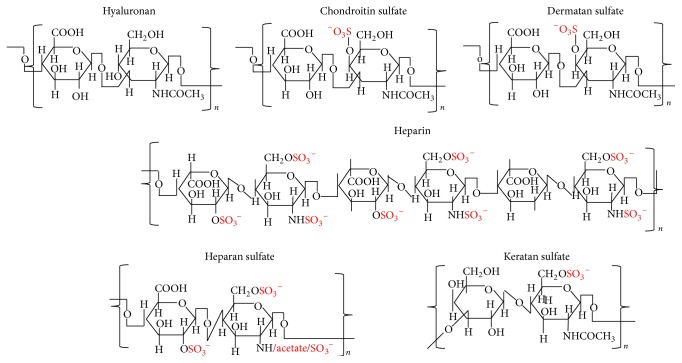
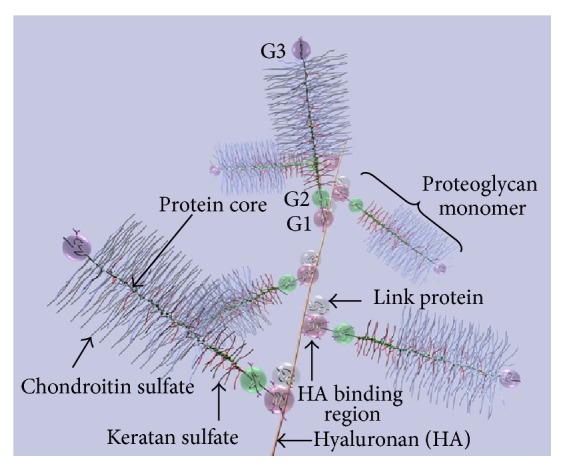
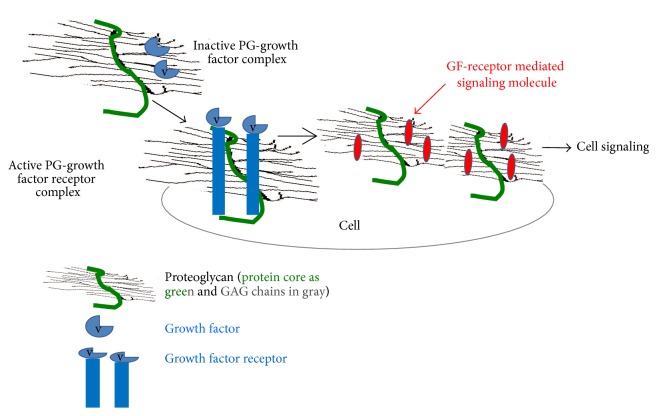
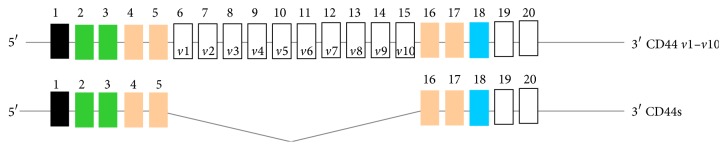
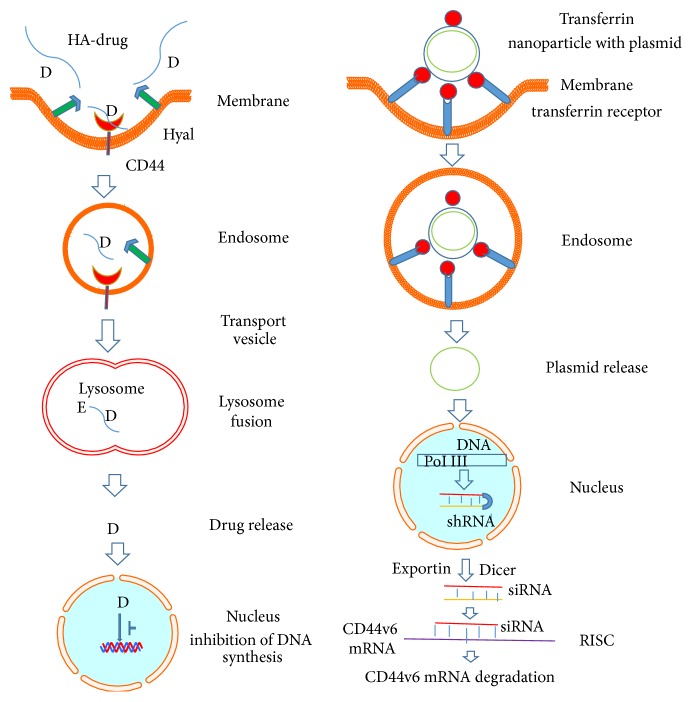
The outcome of patients with cancer has improved significantly in the past decade with the incorporation of drugs targeting cell surface adhesive receptors, receptor tyrosine kinases, and modulation of several molecules of extracellular matrices (ECMs), the complex composite of collagens, glycoproteins, proteoglycans, and glycosaminoglycans that dictates tissue architecture. Cancer tissue invasive processes progress by various oncogenic strategies, including interfering with ECM molecules and their interactions with invasive cells. In this review, we describe how the ECM components, proteoglycans and glycosaminoglycans, influence tumor cell signaling. In particular this review describes how the glycosaminoglycan hyaluronan (HA) and its major receptor CD44 impact invasive behavior of tumor cells, and provides useful insight when designing new therapeutic strategies in the treatment of cancer.
Figures
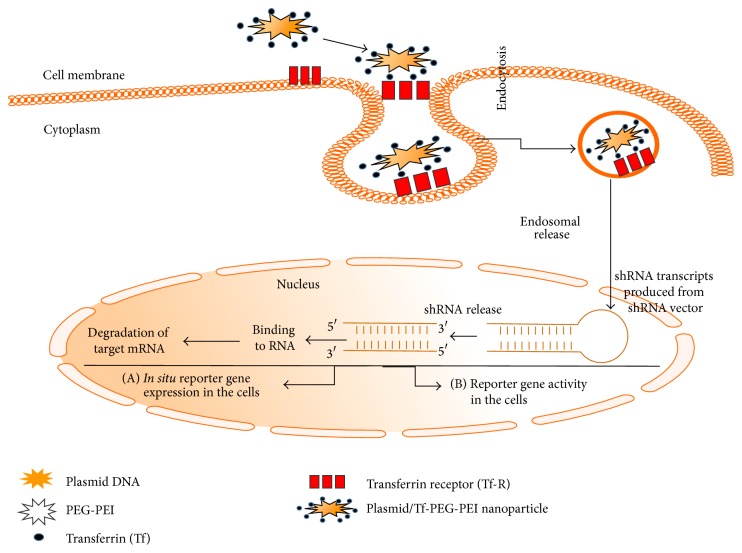
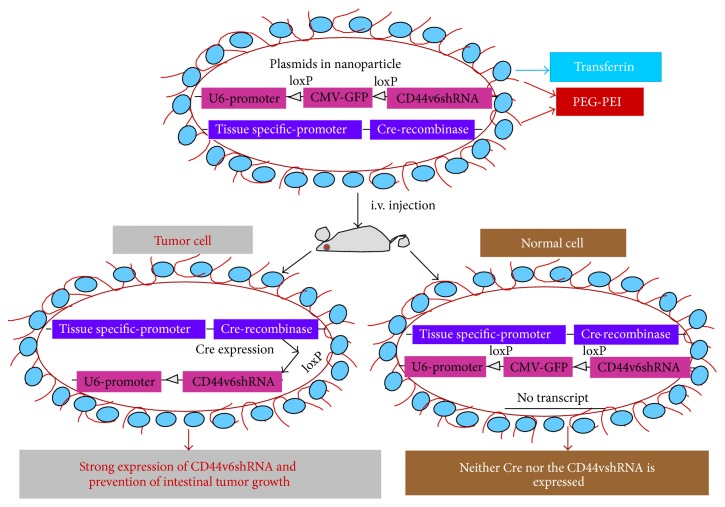
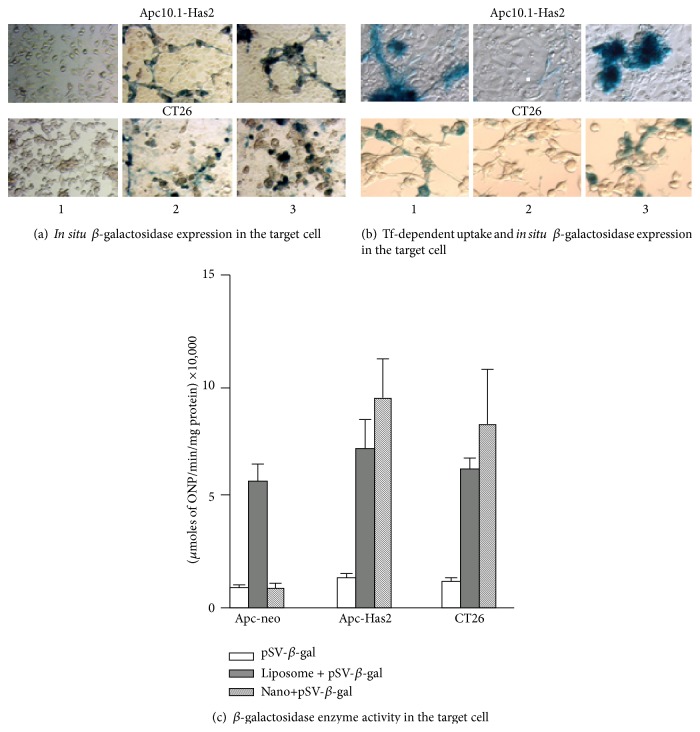
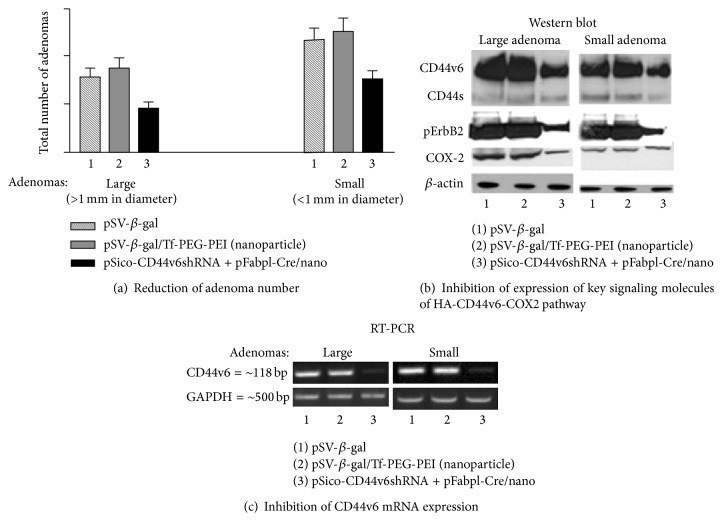
References
-
- Hardingham T. E., Fosang A. J. Proteoglycans: many forms and many functions. The FASEB Journal. 1992;6(3):861–870. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
