

Progressive loss of anti-HER2 CD4+ T-helper type 1 response in breast tumorigenesis and the potential for immune restoration
- PMID: 26451293
- PMCID: PMC4589053
- DOI: 10.1080/2162402X.2015.1022301
Progressive loss of anti-HER2 CD4+ T-helper type 1 response in breast tumorigenesis and the potential for immune restoration
Abstract
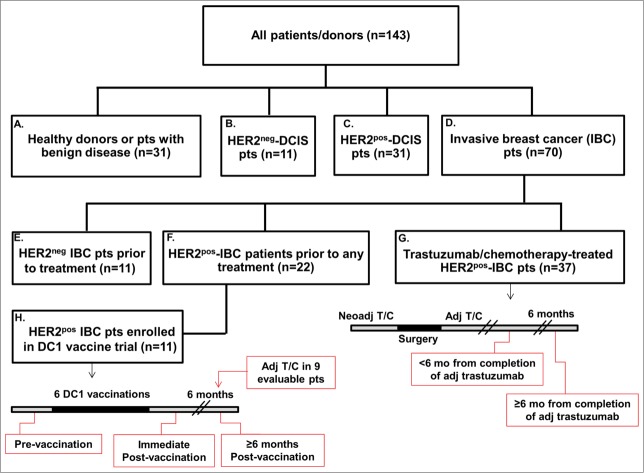
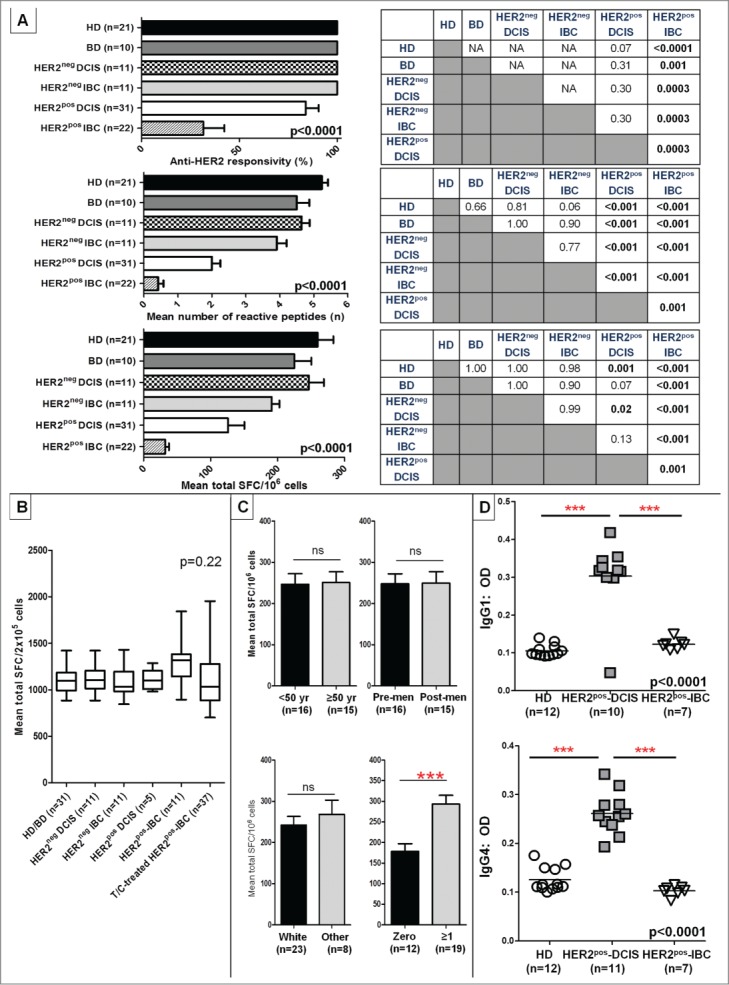
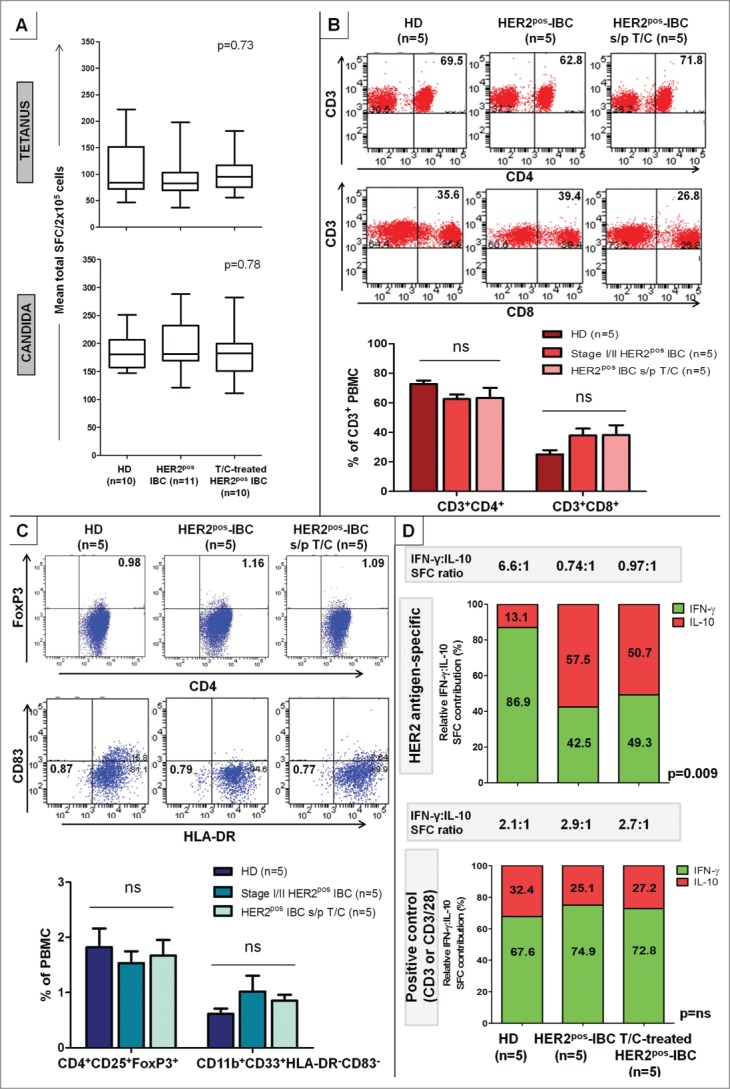
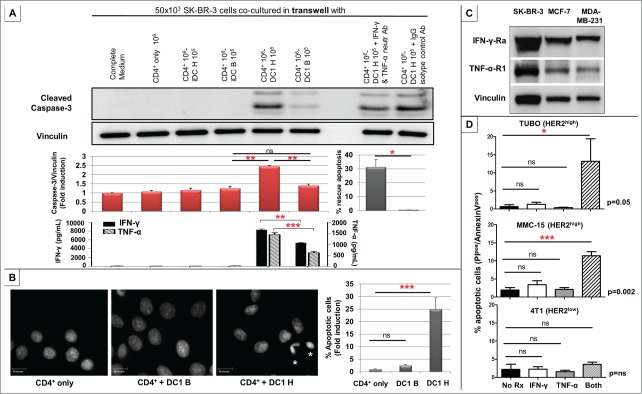
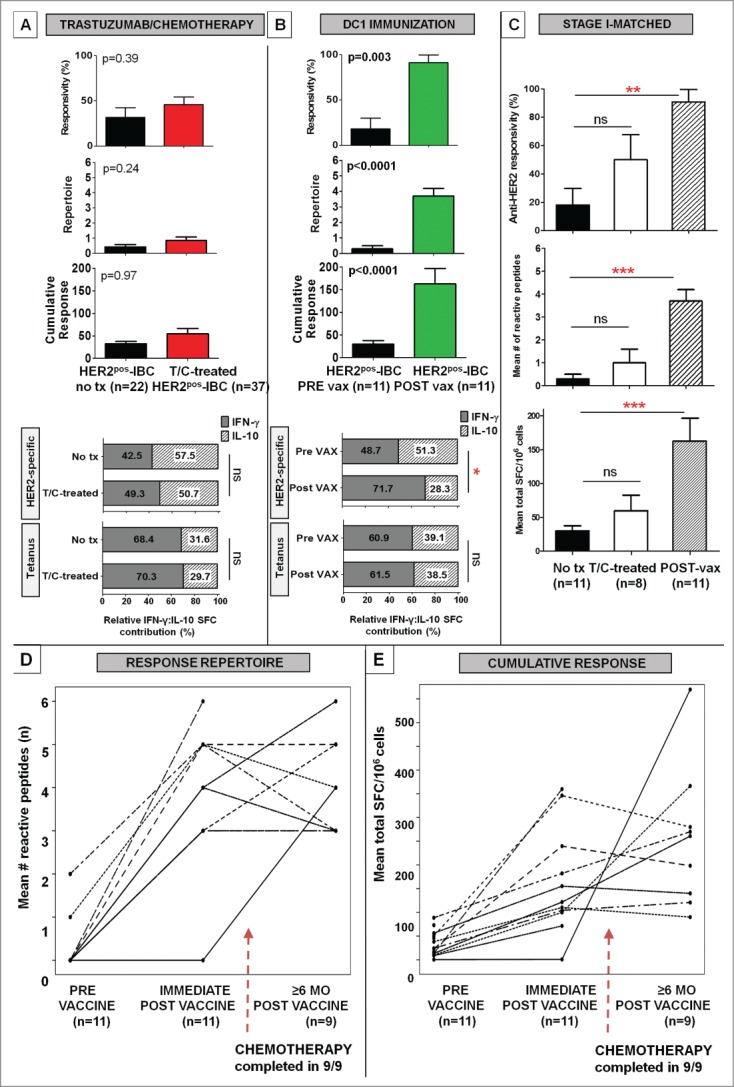
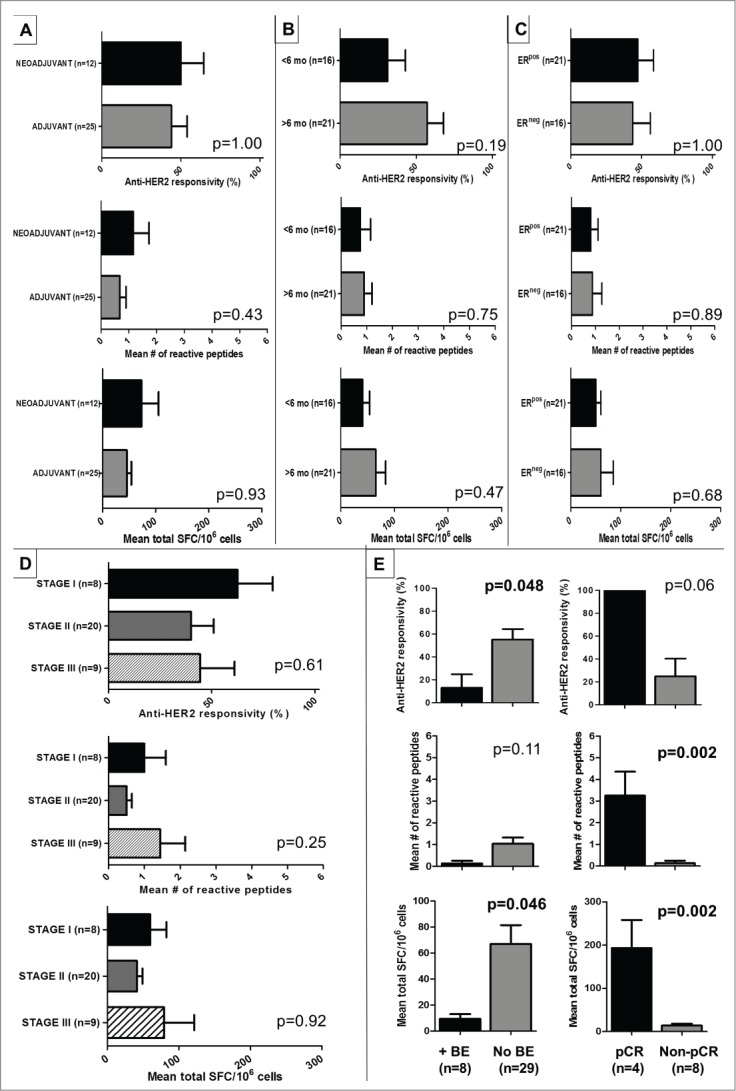
Genomic profiling has identified several molecular oncodrivers in breast tumorigenesis. A thorough understanding of endogenous immune responses to these oncodrivers may provide insights into immune interventions for breast cancer (BC). We investigated systemic anti-HER2/neu CD4+ T-helper type-1 (Th1) responses in HER2-driven breast tumorigenesis. A highly significant stepwise Th1 response loss extending from healthy donors (HD), through HER2pos-DCIS, and ultimately to early stage HER2pos-invasive BC patients was detected by IFNγ ELISPOT. The anti-HER2 Th1 deficit was not attributable to host-level T-cell anergy, loss of immune competence, or increase in immunosuppressive phenotypes (Treg/MDSCs), but rather associated with a functional shift in IFNγ:IL-10-producing phenotypes. HER2high, but not HER2low, BC cells expressing IFNγ/TNF-α receptors were susceptible to Th1 cytokine-mediated apoptosis in vitro, which could be significantly rescued by neutralizing IFNγ and TNF-α, suggesting that abrogation of HER2-specific Th1 may reflect a mechanism of immune evasion in HER2-driven tumorigenesis. While largely unaffected by cytotoxic or HER2-targeted (trastuzumab) therapies, depressed Th1 responses in HER2pos-BC patients were significantly restored following HER2-pulsed dendritic cell (DC) vaccinations, suggesting that this Th1 defect is not "fixed" and can be corrected by immunologic interventions. Importantly, preserved anti-HER2 Th1 responses were associated with pathologic complete response to neoadjuvant trastuzumab/chemotherapy, while depressed responses were observed in patients incurring locoregional/systemic recurrence following trastuzumab/chemotherapy. Monitoring anti-HER2 Th1 reactivity following HER2-directed therapies may identify vulnerable subgroups at risk of clinicopathologic failure. In such patients, combinations of existing HER2-targeted therapies with strategies to boost anti-HER2 CD4+ Th1 immunity may decrease the risk of recurrence and thus warrant further investigation.
Keywords: CD4+ T-helper immunity; HER2/neu; breast cancer; dendritic cell; immune monitoring; immune restoration; vaccination.
Figures
References
-
- Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA Cancer J Clin 2011; 61:69-90; PMID:21296855; http://dx.doi.org/ 10.3322/caac.20107 - DOI - PubMed
-
- Perou CM, Sorlie T, Eisen MB, van de Rijn M, Jeffrey SS, Rees CA, Pollack JR, Ross DT, Johnsen H, Akslen LA et al.. Molecular portraits of human breast tumours. Nature 2000; 406:747-52; PMID:10963602; http://dx.doi.org/ 10.1038/35021093 - DOI - PubMed
-
- Meric F, Hung MC, Hortobagyi GN, Hunt KK. HER2/neu in the management of invasive breast cancer. J Am Coll Surg 2002; 194:488-501; PMID:11949754; http://dx.doi.org/ 10.1016/S1072-7515(02)01121-3 - DOI - PubMed
-
- Henson ES, Hu X, Gibson SB. Herceptin sensitizes ErbB2-overexpressing cells to apoptosis by reducing antiapoptotic Mcl-1 expression. Clin Cancer Res 2006; 12:845-53; PMID:16467098; http://dx.doi.org/ 10.1158/1078-0432.CCR-05-0754 - DOI - PubMed
-
- Wang GS, Zhu H, Bi SJ. Pathological features and prognosis of different molecular subtypes of breast cancer. Mol Med Rep 2012; 6:779-82; PMID:22797840; http://dx.doi.org/ 10.3892/mmr.2012.981 - DOI - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous