

A transposable element insertion in the susceptibility gene CsaMLO8 results in hypocotyl resistance to powdery mildew in cucumber
- PMID: 26453551
- PMCID: PMC4600303
- DOI: 10.1186/s12870-015-0635-x
A transposable element insertion in the susceptibility gene CsaMLO8 results in hypocotyl resistance to powdery mildew in cucumber
Abstract
Background: Powdery mildew (PM) is an important disease of cucumber (Cucumis sativus L.). CsaMLO8 was previously identified as a candidate susceptibility gene for PM in cucumber, for two reasons: 1) This gene clusters phylogenetically in clade V, which has previously been shown to harbour all known MLO-like susceptibility genes for PM identified in dicot species; 2) This gene co-localizes with a QTL on chromosome 5 for hypocotyl-specific resistance to PM.
Methods: CsaMLO8 alleles from susceptible and resistant cucumber were cloned and transformed to mlo-mutant tomato. Cucumber seedlings were inoculated with Podosphaera xanthii, tissues were studied for CsaMLO8 expression at several timepoints post inoculation using qRT-PCR. The occurrence of the observed loss-of-function allele of CsaMLO8 in resequenced cucumber accessions was studied in silico.
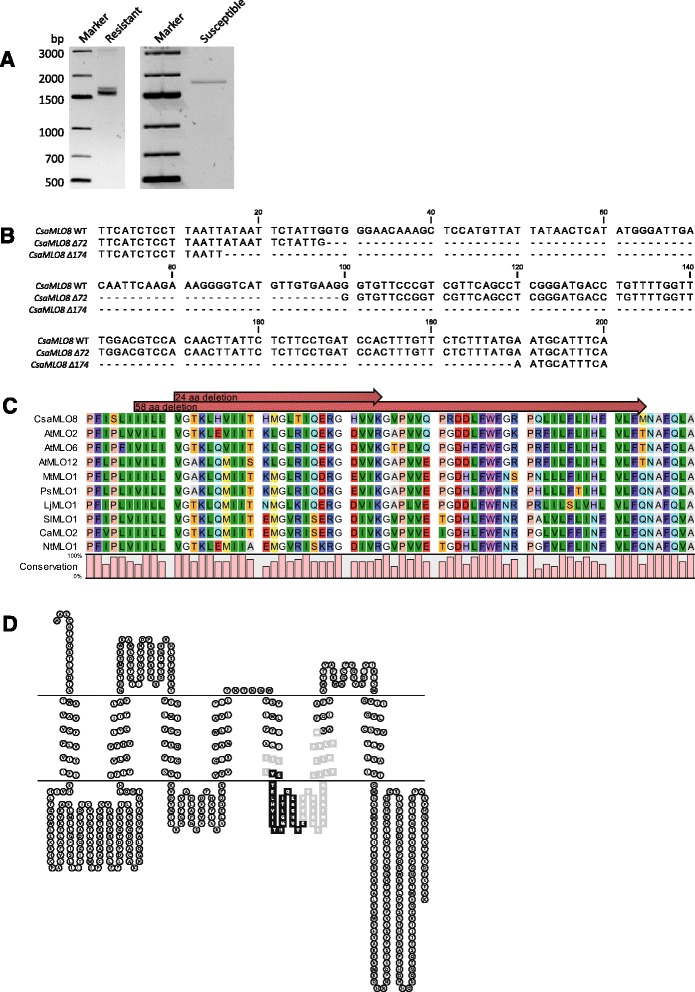
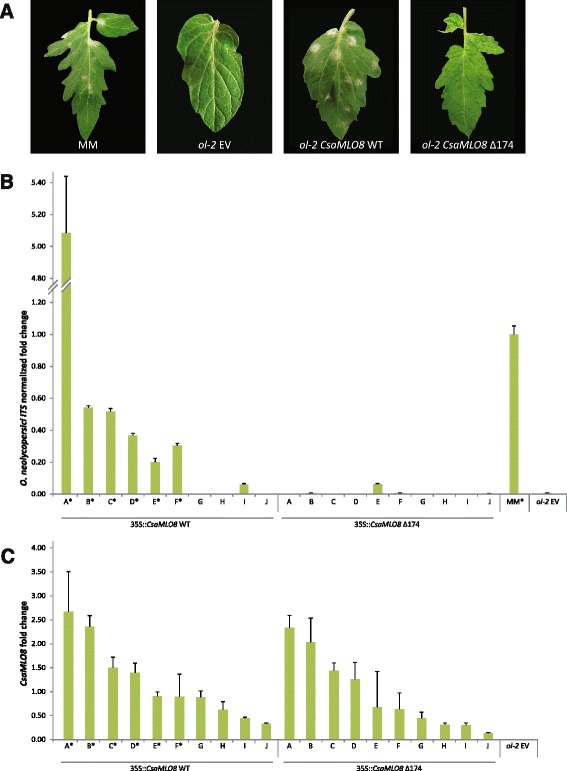
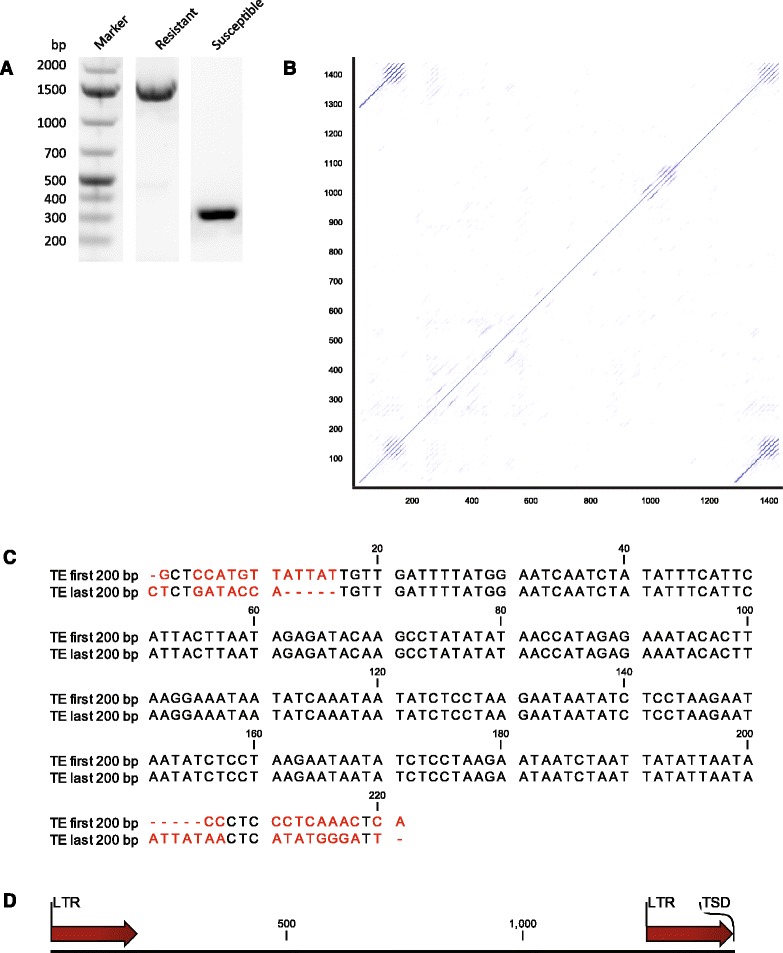
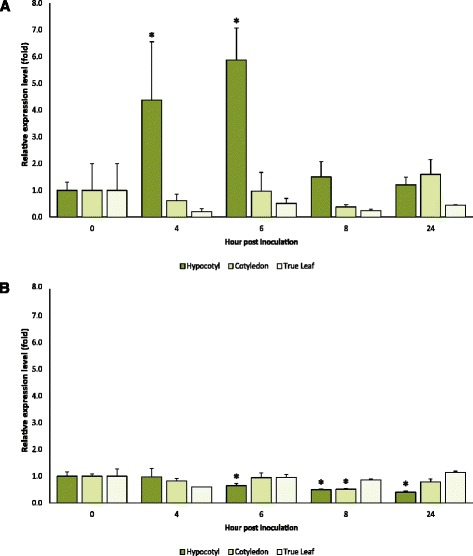
Results: We cloned CsaMLO8 alleles from susceptible and resistant cucumber genotypes, the latter carrying the QTL for hypocotyl resistance. We found that insertion of a non-autonomous Class LTR retrotransposable element in the resistant genotype leads to aberrant splicing of CsaMLO8 mRNA. Heterologous expression of the wild-type allele of CsaMLO8 in a tomato mlo-mutant restored PM susceptibility. However, heterologous expression of the CsaMLO8 allele cloned from the resistant cucumber genotype failed to restore PM susceptibility. Furthermore we showed that inoculation of susceptible cucumber with the PM pathogen Podosphaera xanthii induced transcriptional upregulation of CsaMLO8 in hypocotyl tissue, but not in cotyledon or leaf tissue. This coincides with the observation that the QTL at the CsaMLO8-locus causes full resistance in hypocotyl tissue, but only partial resistance in cotyledons and true leafs. We studied the occurrence of the loss-of-function allele of CsaMLO8 in cucumber germplasm by an in silico approach using resequencing data of a collection of 115 cucumber accessions, and found that this allele was present in 31 out of 115 accessions.
Conclusions: CsaMLO8 was characterised as a functional susceptibility gene to PM, particularly in the hypocotyl where it was transcriptionally upregulated upon inoculation with the PM pathogen P. xanthii. A loss-of-function mutation in CsaMLO8 due to the insertion of a transposable element was found to be the cause of hypocotyl resistance to PM. This particular allele of CsaMLO8 was found to occur in 27 % of the resequenced cucumber accessions.
Figures
References
-
- Food and Agriculture Organization of the United Nations, Statistics Division. [http://faostat3.fao.org/] Accessed May 2015.
-
- Block CC, Reitsma KR. Powdery Mildew Resistance in the U. S. National Plant Germplasm System Cucumber Collection. Hortic Sci. 2005;40:416–20.
-
- Jahn M, Munger HM, McCreight JD. The powdery mildews: a comprehensive treatise. 2002. Breeding cucurbit crops for powdery mildew resistance; pp. 239–48.
-
- Shanmugasundaram S, Williams PH, Peterson CE. Inheritance of resistance to powdery mildew in cucumber. Phytopathology. 1971;61:1218–21. doi: 10.1094/Phyto-61-1218. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
