

Deciphering Genome Content and Evolutionary Relationships of Isolates from the Fungus Magnaporthe oryzae Attacking Different Host Plants
- PMID: 26454013
- PMCID: PMC4684704
- DOI: 10.1093/gbe/evv187
Deciphering Genome Content and Evolutionary Relationships of Isolates from the Fungus Magnaporthe oryzae Attacking Different Host Plants
Abstract
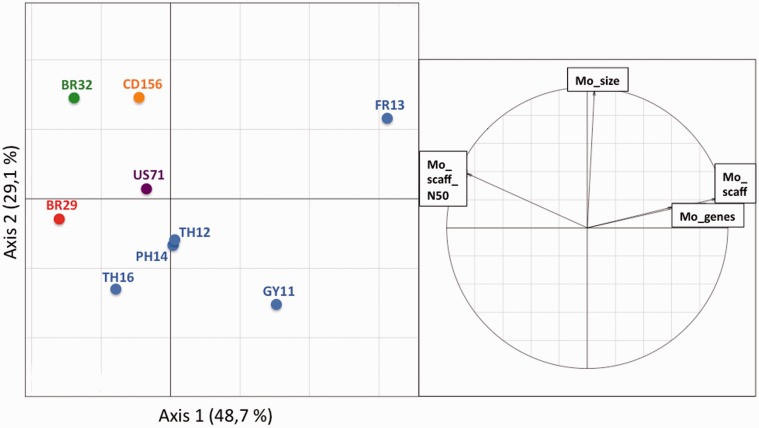
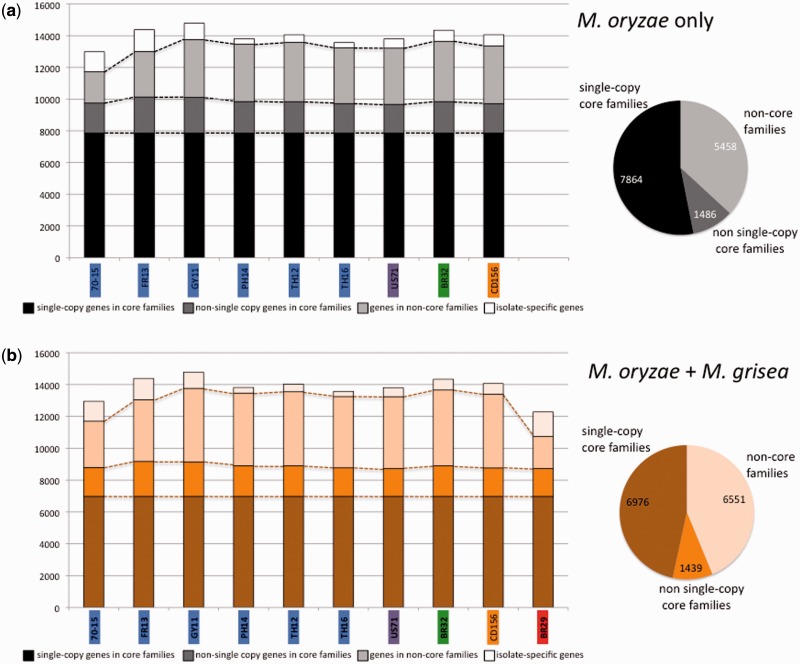
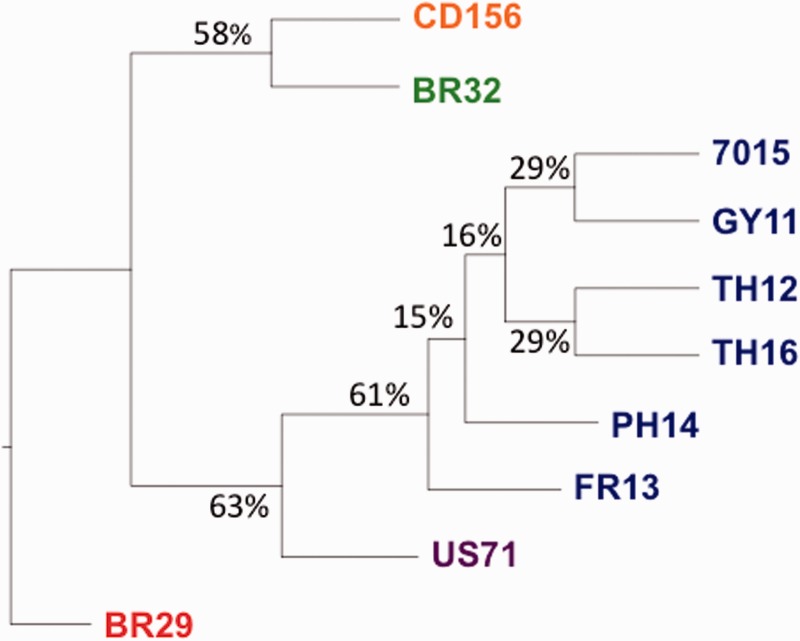
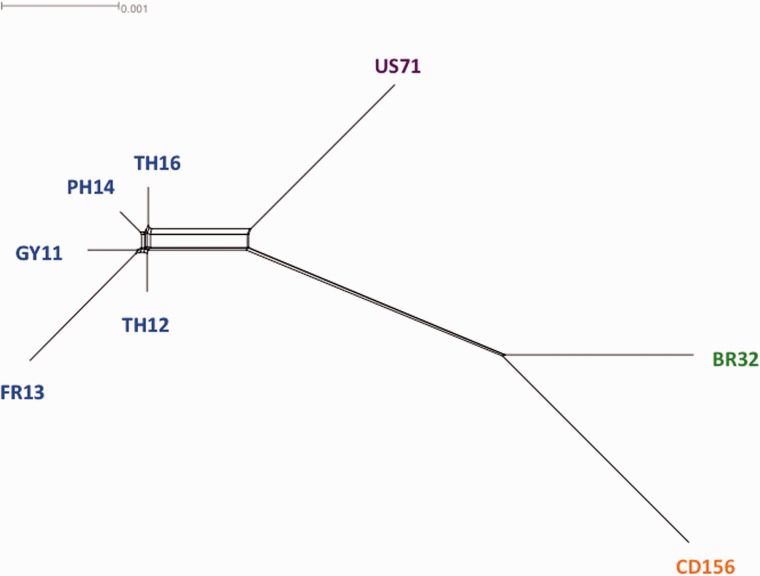
Deciphering the genetic bases of pathogen adaptation to its host is a key question in ecology and evolution. To understand how the fungus Magnaporthe oryzae adapts to different plants, we sequenced eight M. oryzae isolates differing in host specificity (rice, foxtail millet, wheat, and goosegrass), and one Magnaporthe grisea isolate specific of crabgrass. Analysis of Magnaporthe genomes revealed small variation in genome sizes (39-43 Mb) and gene content (12,283-14,781 genes) between isolates. The whole set of Magnaporthe genes comprised 14,966 shared families, 63% of which included genes present in all the nine M. oryzae genomes. The evolutionary relationships among Magnaporthe isolates were inferred using 6,878 single-copy orthologs. The resulting genealogy was mostly bifurcating among the different host-specific lineages, but was reticulate inside the rice lineage. We detected traces of introgression from a nonrice genome in the rice reference 70-15 genome. Among M. oryzae isolates and host-specific lineages, the genome composition in terms of frequencies of genes putatively involved in pathogenicity (effectors, secondary metabolism, cazome) was conserved. However, 529 shared families were found only in nonrice lineages, whereas the rice lineage possessed 86 specific families absent from the nonrice genomes. Our results confirmed that the host specificity of M. oryzae isolates was associated with a divergence between lineages without major gene flow and that, despite the strong conservation of gene families between lineages, adaptation to different hosts, especially to rice, was associated with the presence of a small number of specific gene families. All information was gathered in a public database (http://genome.jouy.inra.fr/gemo).
Keywords: adaptation to the host, rice blast, comparative genomics; http://genome.jouy.inra.fr/gemo.
© The Author(s) 2015. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures
References
-
- Aguileta G, et al. 2010. Finding candidate genes under positive selection in non-model species: examples of genes involved in host specialization in pathogens. Mol Ecol. 19:292–306. - PubMed
-
- Anderson PK, et al. 2004. Emerging infectious diseases of plants: pathogen pollution, climate change and agrotechnology drivers. Trends Ecol Evol. 19:535–544. - PubMed
-
- Ané C, Larget B, Baum DA, Smith SD, Rokas A. 2007. Bayesian estimation of concordance among gene trees. Mol Biol Evol. 24:412–426. - PubMed
Publication types
MeSH terms
Substances
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
