

Elevated CO2-Induced Responses in Stomata Require ABA and ABA Signaling
- PMID: 26455301
- PMCID: PMC4612465
- DOI: 10.1016/j.cub.2015.09.013
Elevated CO2-Induced Responses in Stomata Require ABA and ABA Signaling
Abstract
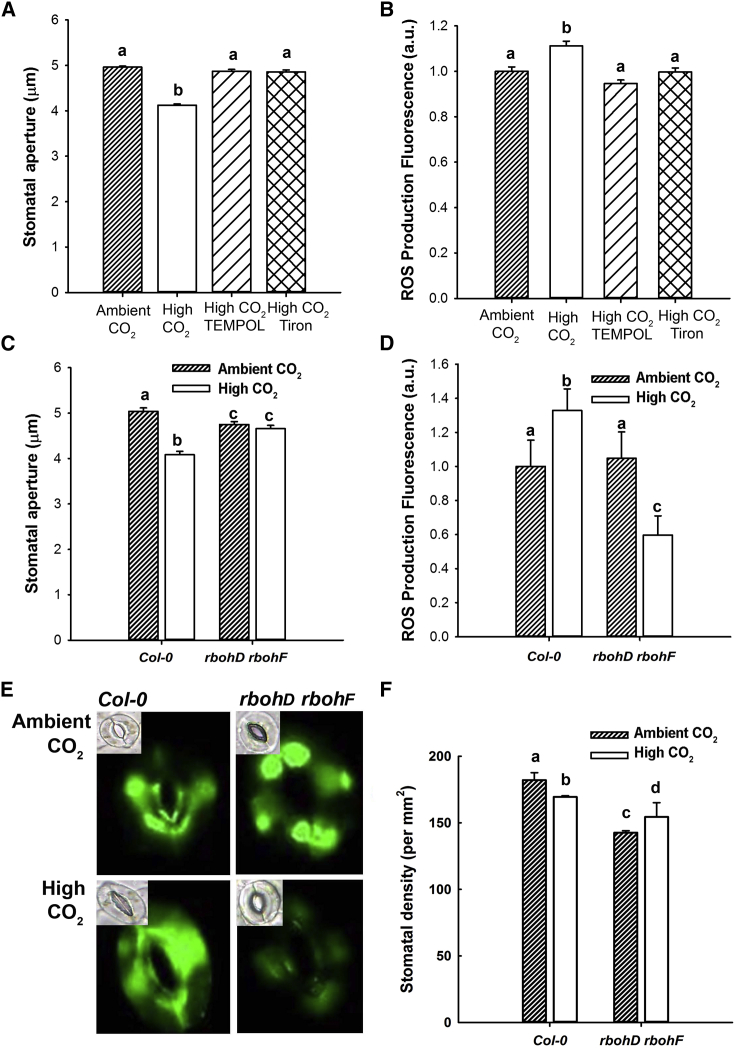
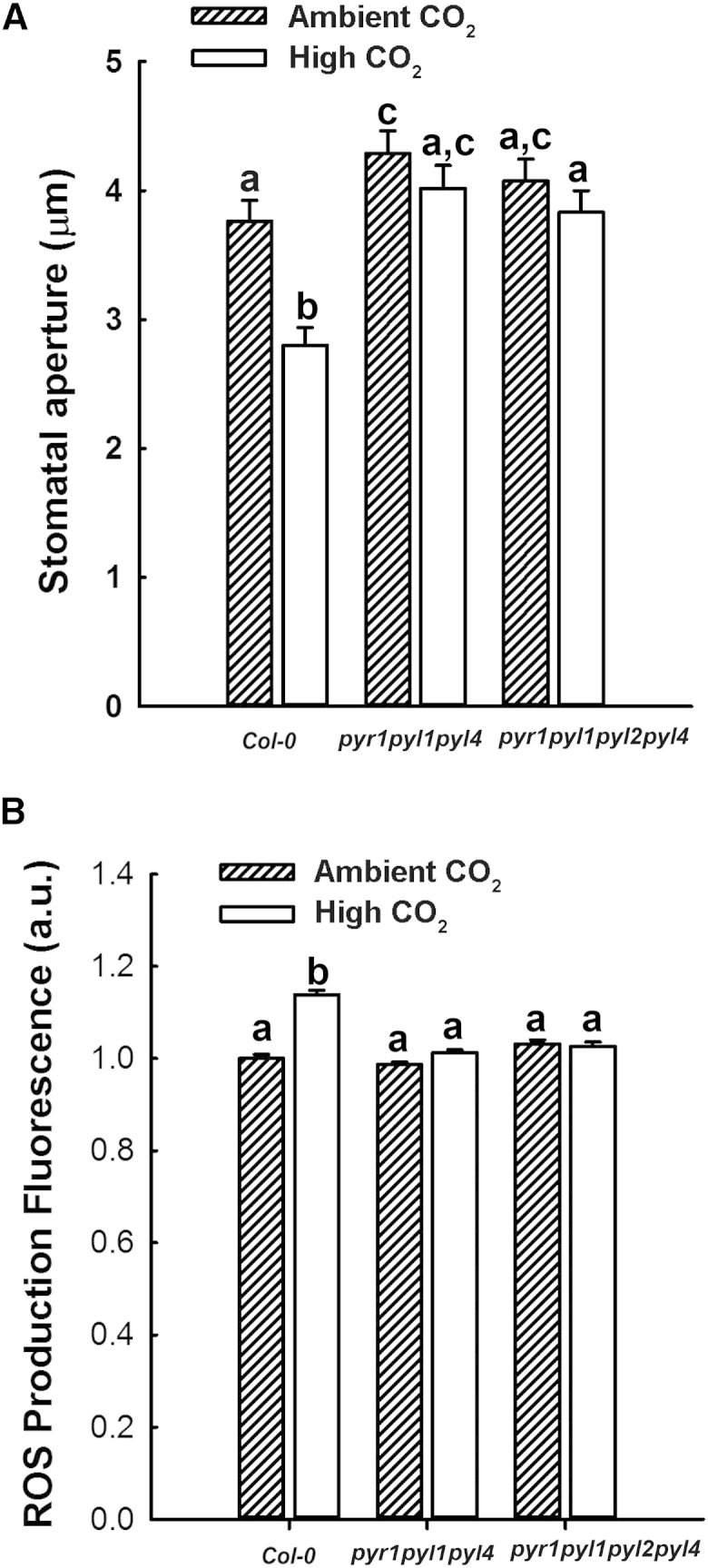
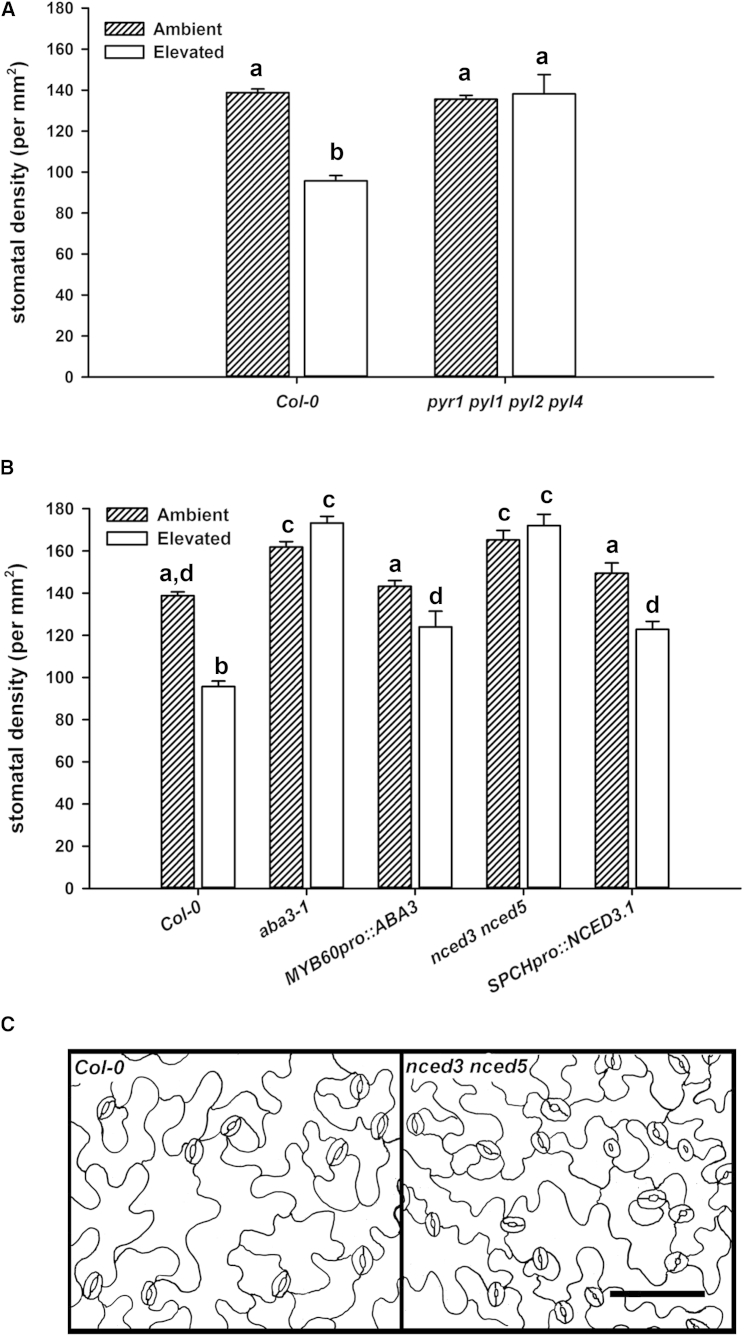
An integral part of global environment change is an increase in the atmospheric concentration of CO2 ([CO2]) [1]. Increased [CO2] reduces leaf stomatal apertures and density of stomata that plays out as reductions in evapotranspiration [2-4]. Surprisingly, given the importance of transpiration to the control of terrestrial water fluxes [5] and plant nutrient acquisition [6], we know comparatively little about the molecular components involved in the intracellular signaling pathways by which [CO2] controls stomatal development and function [7]. Here, we report that elevated [CO2]-induced closure and reductions in stomatal density require the generation of reactive oxygen species (ROS), thereby adding a new common element to these signaling pathways. We also show that the PYR/RCAR family of ABA receptors [8, 9] and ABA itself are required in both responses. Using genetic approaches, we show that ABA in guard cells or their precursors is sufficient to mediate the [CO2]-induced stomatal density response. Taken together, our results suggest that stomatal responses to increased [CO2] operate through the intermediacy of ABA. In the case of [CO2]-induced reductions in stomatal aperture, this occurs by accessing the guard cell ABA signaling pathway. In both [CO2]-mediated responses, our data are consistent with a mechanism in which ABA increases the sensitivity of the system to [CO2] but could also be explained by requirement for a CO2-induced increase in ABA biosynthesis specifically in the guard cell lineage. Furthermore, the dependency of stomatal [CO2] signaling on ABA suggests that the ABA pathway is, in evolutionary terms, likely to be ancestral.
Keywords: ABA receptors; ABA signaling; NADPH oxidases; ROS; Rboh genes; [CO(2)] signaling; guard cells; signaling convergence; stomata; stomatal closure; stomatal density.
Copyright © 2015 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures


Similar articles
-
Abscisic acid-independent stomatal CO2 signal transduction pathway and convergence of CO2 and ABA signaling downstream of OST1 kinase.Proc Natl Acad Sci U S A. 2018 Oct 16;115(42):E9971-E9980. doi: 10.1073/pnas.1809204115. Epub 2018 Oct 3. Proc Natl Acad Sci U S A. 2018. PMID: 30282744 Free PMC article.
-
PYR/RCAR receptors contribute to ozone-, reduced air humidity-, darkness-, and CO2-induced stomatal regulation.Plant Physiol. 2013 Jul;162(3):1652-68. doi: 10.1104/pp.113.220608. Epub 2013 May 23. Plant Physiol. 2013. PMID: 23703845 Free PMC article.
-
Ethylene Inhibits Methyl Jasmonate-Induced Stomatal Closure by Modulating Guard Cell Slow-Type Anion Channel Activity via the OPEN STOMATA 1/SnRK2.6 Kinase-Independent Pathway in Arabidopsis.Plant Cell Physiol. 2019 Oct 1;60(10):2263-2271. doi: 10.1093/pcp/pcz121. Plant Cell Physiol. 2019. PMID: 31241163
-
Signaling Transduction of ABA, ROS, and Ca2+ in Plant Stomatal Closure in Response to Drought.Int J Mol Sci. 2022 Nov 26;23(23):14824. doi: 10.3390/ijms232314824. Int J Mol Sci. 2022. PMID: 36499153 Free PMC article. Review.
-
A brand new START: abscisic acid perception and transduction in the guard cell.Sci Signal. 2011 Nov 29;4(201):re4. doi: 10.1126/scisignal.2002164. Sci Signal. 2011. PMID: 22126965 Review.
Cited by
-
Stomatal development in the changing climate.Development. 2024 Oct 15;151(20):dev202681. doi: 10.1242/dev.202681. Epub 2024 Oct 21. Development. 2024. PMID: 39431330 Free PMC article. Review.
-
Effect of CO2 Elevation on Tomato Gas Exchange, Root Morphology and Water Use Efficiency under Two N-Fertigation Levels.Plants (Basel). 2024 Aug 26;13(17):2373. doi: 10.3390/plants13172373. Plants (Basel). 2024. PMID: 39273857 Free PMC article.
-
Climate change shapes the future evolution of plant metabolism.Adv Genet (Hoboken). 2020 Aug 10;1(1):e10022. doi: 10.1002/ggn2.10022. eCollection 2020 Dec. Adv Genet (Hoboken). 2020. PMID: 36619247 Free PMC article.
-
Barley Genotypes Vary in Stomatal Responsiveness to Light and CO2 Conditions.Plants (Basel). 2021 Nov 21;10(11):2533. doi: 10.3390/plants10112533. Plants (Basel). 2021. PMID: 34834896 Free PMC article.
-
Signaling mechanisms in abscisic acid-mediated stomatal closure.Plant J. 2021 Jan;105(2):307-321. doi: 10.1111/tpj.15067. Epub 2020 Dec 9. Plant J. 2021. PMID: 33145840 Free PMC article. Review.
References
-
- Long S.P., Ainsworth E.A., Rogers A., Ort D.R. Rising atmospheric carbon dioxide: plants FACE the future. Annu. Rev. Plant Biol. 2004;55:591–628. - PubMed
-
- Mansfield T.A., Hetherington A.M., Atkinson C.J. Some current aspects of stomatal physiology. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1990;41:55–75.
-
- Vavasseur A., Raghavendra A.S. Guard cell metabolism and CO2 sensing. New Phytol. 2005;165:665–682. - PubMed
-
- Jasechko S., Sharp Z.D., Gibson J.J., Birks S.J., Yi Y., Fawcett P.J. Terrestrial water fluxes dominated by transpiration. Nature. 2013;496:347–350. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
