

Natural Variation in plep-1 Causes Male-Male Copulatory Behavior in C. elegans
- PMID: 26455306
- PMCID: PMC4618159
- DOI: 10.1016/j.cub.2015.09.019
Natural Variation in plep-1 Causes Male-Male Copulatory Behavior in C. elegans
Abstract
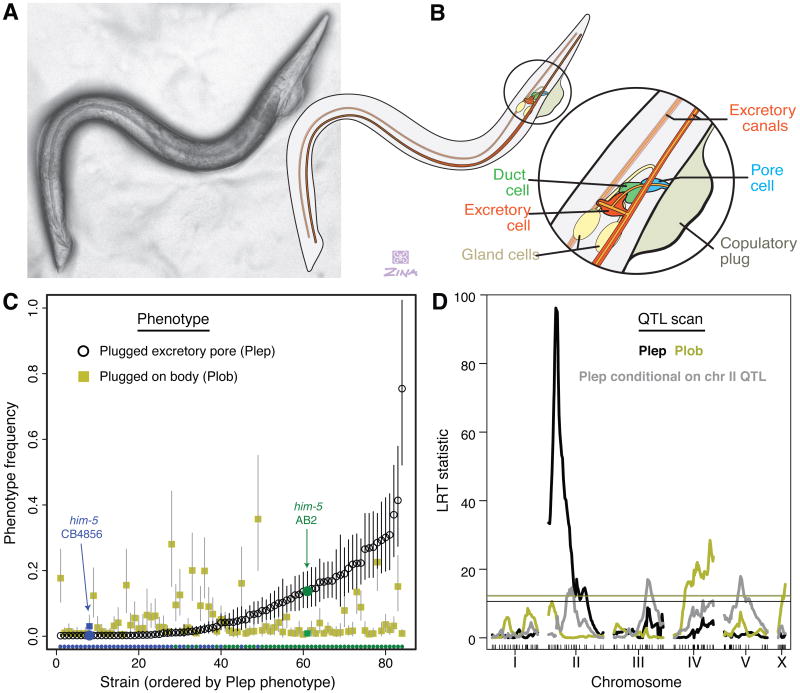
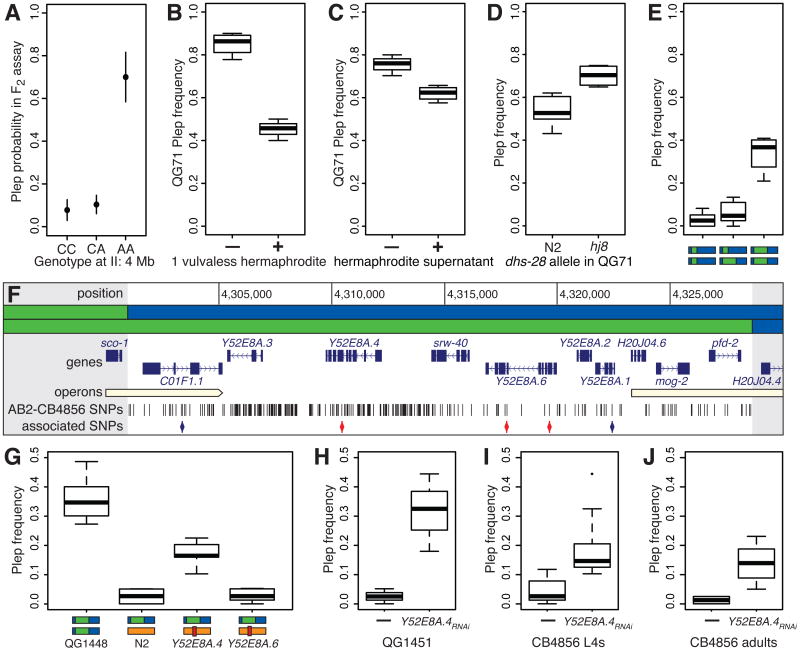
In sexual species, gametes have to find and recognize one another. Signaling is thus central to sexual reproduction and involves a rapidly evolving interplay of shared and divergent interests [1-4]. Among Caenorhabditis nematodes, three species have evolved self-fertilization, changing the balance of intersexual relations [5]. Males in these androdioecious species are rare, and the evolutionary interests of hermaphrodites dominate. Signaling has shifted accordingly, with females losing behavioral responses to males [6, 7] and males losing competitive abilities [8, 9]. Males in these species also show variable same-sex and autocopulatory mating behaviors [6, 10]. These behaviors could have evolved by relaxed selection on male function, accumulation of sexually antagonistic alleles that benefit hermaphrodites and harm males [5, 11], or neither of these, because androdioecy also reduces the ability of populations to respond to selection [12-14]. We have identified the genetic cause of a male-male mating behavior exhibited by geographically dispersed C. elegans isolates, wherein males mate with and deposit copulatory plugs on one another's excretory pores. We find a single locus of major effect that is explained by segregation of a loss-of-function mutation in an uncharacterized gene, plep-1, expressed in the excretory cell in both sexes. Males homozygous for the plep-1 mutation have excretory pores that are attractive or receptive to copulatory behavior of other males. Excretory pore plugs are injurious and hermaphrodite activity is compromised in plep-1 mutants, so the allele might be unconditionally deleterious, persisting in the population because the species' androdioecious mating system limits the reach of selection.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures
Comment in
-
Reproductive Evolution: Pulling the Plug on Selection.Curr Biol. 2015 Oct 19;25(20):R984-6. doi: 10.1016/j.cub.2015.09.008. Curr Biol. 2015. PMID: 26485371
Similar articles
-
Diversity in mating behavior of hermaphroditic and male-female Caenorhabditis nematodes.Genetics. 2007 Apr;175(4):1761-71. doi: 10.1534/genetics.106.068304. Epub 2007 Feb 4. Genetics. 2007. PMID: 17277358 Free PMC article.
-
Molecular basis of the copulatory plug polymorphism in Caenorhabditis elegans.Nature. 2008 Aug 21;454(7207):1019-22. doi: 10.1038/nature07171. Epub 2008 Jul 16. Nature. 2008. PMID: 18633349 Free PMC article.
-
When females produce sperm: genetics of C. elegans hermaphrodite reproductive choice.G3 (Bethesda). 2013 Oct 3;3(10):1851-9. doi: 10.1534/g3.113.007914. G3 (Bethesda). 2013. PMID: 23979940 Free PMC article.
-
The evolution from females to hermaphrodites results in a sexual conflict over mating in androdioecious nematode worms and clam shrimp.J Evol Biol. 2010 Mar;23(3):539-56. doi: 10.1111/j.1420-9101.2009.01919.x. Epub 2010 Jan 14. J Evol Biol. 2010. PMID: 20074309 Review.
-
Males, Outcrossing, and Sexual Selection in Caenorhabditis Nematodes.Genetics. 2019 Sep;213(1):27-57. doi: 10.1534/genetics.119.300244. Genetics. 2019. PMID: 31488593 Free PMC article. Review.
Cited by
-
Natural Variation and Genetic Determinants of Caenorhabditis elegans Sperm Size.Genetics. 2019 Oct;213(2):615-632. doi: 10.1534/genetics.119.302462. Epub 2019 Aug 8. Genetics. 2019. PMID: 31395653 Free PMC article.
-
Natural sensory context drives diverse brain-wide activity during C. elegans mating.Cell. 2021 Sep 30;184(20):5122-5137.e17. doi: 10.1016/j.cell.2021.08.024. Epub 2021 Sep 16. Cell. 2021. PMID: 34534446 Free PMC article.
-
Polygenicity and Epistasis Underlie Fitness-Proximal Traits in the Caenorhabditis elegans Multiparental Experimental Evolution (CeMEE) Panel.Genetics. 2017 Dec;207(4):1663-1685. doi: 10.1534/genetics.117.300406. Epub 2017 Oct 24. Genetics. 2017. PMID: 29066469 Free PMC article.
-
From QTL to gene: C. elegans facilitates discoveries of the genetic mechanisms underlying natural variation.Trends Genet. 2021 Oct;37(10):933-947. doi: 10.1016/j.tig.2021.06.005. Epub 2021 Jul 3. Trends Genet. 2021. PMID: 34229867 Free PMC article. Review.
-
Sexual Dimorphism and Sex Differences in Caenorhabditis elegans Neuronal Development and Behavior.Genetics. 2018 Mar;208(3):909-935. doi: 10.1534/genetics.117.300294. Genetics. 2018. PMID: 29487147 Free PMC article.
References
-
- Smith JM, Harper D. Animal Signals. Oxford University Press; 2003.
-
- Halliday TR. The study of mate choice. In: Bateson P, editor. Mate Choice. University of Cambridge; 1983. pp. 3–32.
-
- Johansson BG, Jones TM. The role of chemical communication in mate choice. Biological Reviews. 2007;82:265–289. - PubMed
-
- Wyatt TD. Pheromones and Animal Behaviour. Cambridge University Press; 2003.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
